




















 biology
biologySimilar presentations:




")

")



Эмбриология. Половые клетки, оплодотворение. Дробление
1.
Занятие №5Тема: Эмбриология. Половые клетки, Оплодотворение.Дробление
Цели и задачи:
1.Познакомиться со строением сперматозоидов.
2.Познакомиться с процессом сперматогенеза.
3. Познакомиться с гормональной регуляцией процесса сперматогенеза.
4.Познакомиться с процессом образования женских половых клеток
5. Познакомиться со строением яйцеклетки.
6. Познакомиться с гормональной регуляцией овогенеза.
7. Познакомиться с процессом оплодотворения.
Задание для самостоятельной работы студентов
1.Зарисовать слайды: 11,14,19(2),20(2),25.
Состальными слайдами ознакомиться.
2. Ответить на контрольные вопросы.
3. Решить тестовые задания.
2.
Слайд 1ТЕРМИНЫ К ЗАНЯТИЮ
Эмбриология – это наука, изучающая образование половых клеток, их строение, оплодотворение, а
также закономерности развития зародыша.
Гаме́ты (от греч. γᾰμετή — жена, γᾰμέτης — муж), или половые клетки, — репродуктивные клетки,
имеющие гаплоидный (одинарный) набор хромосом и участвующие в гаметном, в частности половом,
размножении.
Эякулят – порция выделеного семени.
Прогенез - это период развития и созревания половых клеток – гамет
(яйцеклеток и сперматозоидов) В результате прогенеза в зрелых половых клетках возникает
гаплоидный набор хромосом, а также формируются структуры обеспечивающие способность к
оплодотворению и развитию зародыша.
Акросома – плоский мешочек, покрывающий переднюю часть ядра головки сперматозоида
Спе́рма семя, эякулят — жидкость (мутная, вязкая, опалесцирующая, светло-серого цвета),
выделяемая приэякуляции (семяизвержении) самцами животных, Сперма – смесь спермfnjpjbljd
(половых клеток самца) и плазмы (сыворотки).
Сперматозоид — это мужская половая клетка (гамета)
3.
Слайд 2ТЕРМИНЫ К ЗАНЯТИЮ
ПОЛОЖИТЕЛЬНЫЙ РЕОТАКСИС – движение сперматозоида против тока жидкости, выделяемой маточными
трубами
ХЕМОТКСИС – движение в сторону химического соединения.
половые клетки выделяют гормоны (гамоны); ♀ - гиногамоны, у ♂ - андрогамоны.
КАПОЦИТАЦИЯ – приобретение сперматозоидом оплодотворяющей способности под действием секрета
маточных труб.
КОНТАКТНОЕ ВЗАИМОДЕЙСТВИЕ – сперматозоид раздвигает лучистый венец и образует канал в блестящей
оболочке. Сперматозоид контактирует с рецептором (Zp3 белок) и начинается акросомная реакция – экзоцитоз
содержимого акросомы для локального разрушения прозрачной оболочки. Внутри головки происходит: внутрь
проникают ионы Ca и Na, наружу H2. Результат – увеличение концентрации Ca запускает ряд процессов, ведущих
к увеличению внутриклеточного pH, запускает акросомную реакцию. Результат акросомной реакции –
образование канала в прозрачной оболочке, через который проходит сперматозоид. В оолемму встраивается
мембрана сперматозоида..
АКТИВАЦИЯ ЯЙЦЕКЛЕТКИ – участок мембраны яйцеклетки, полученный от сперматозоида проницаем для
ионов Na→изменяется потенциа клетки→ионы Ca выходят из клеточного депо в цитоплазму→экзоцитоз
кортикальных гранул. Это приводит к образованию оболочки оплодотворения – изменение свойств блестящей
оболочки, она препятствует полиспермии.
4.
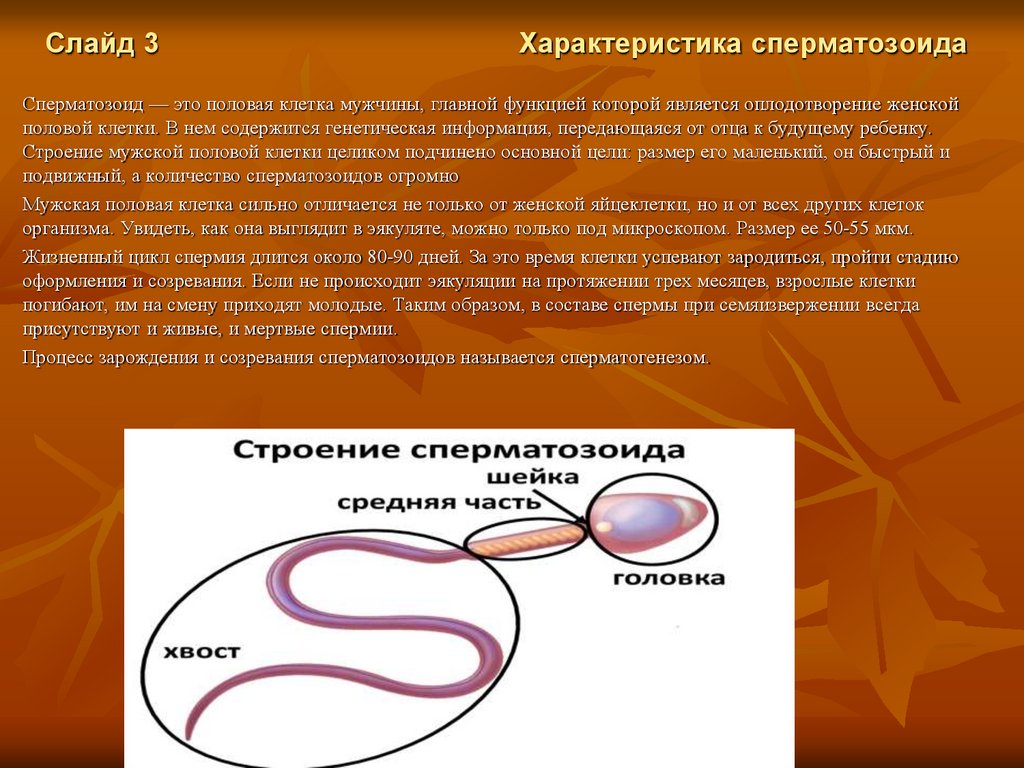
Слайд 3Характеристика сперматозоида
Сперматозоид — это половая клетка мужчины, главной функцией которой является оплодотворение женской
половой клетки. В нем содержится генетическая информация, передающаяся от отца к будущему ребенку.
Строение мужской половой клетки целиком подчинено основной цели: размер его маленький, он быстрый и
подвижный, а количество сперматозоидов огромно
Мужская половая клетка сильно отличается не только от женской яйцеклетки, но и от всех других клеток
организма. Увидеть, как она выглядит в эякуляте, можно только под микроскопом. Размер ее 50-55 мкм.
Жизненный цикл спермия длится около 80-90 дней. За это время клетки успевают зародиться, пройти стадию
оформления и созревания. Если не происходит эякуляции на протяжении трех месяцев, взрослые клетки
погибают, им на смену приходят молодые. Таким образом, в составе спермы при семяизвержении всегда
присутствуют и живые, и мертвые спермии.
Процесс зарождения и созревания сперматозоидов называется сперматогенезом.
5.
Слайд 4Строение сперматозоида
Сперматозоиды — высокоспециализированные клетки, не способные к росту и делению. Сперматозоид состоит
из головки, в которой находится отцовский генетический материал (ДНК), и хвоста, обеспечивающего его
подвижность. При наличии большого ядра у сперматозоида практически нет цитоплазмы, присутствие которой
характерно для большинства клеток организма. Морфологическая гетерогенность эякулята каждого мужчины
уникальна.
Головка сперматозоидов. Она имеет вид эллипсоида, с незначительными вмятинами с обоих боков. С одной из
сторон спермия присутствует также маленькое углубление. Именно благодаря ему головку сперматозоида
называют «ложковидной».
Головки окрашенных сперматозоидов несколько меньше по размерам, чем головки живых сперматозоидов в
сперме. В норме головка имеет овальную форму, длину около 4,0-5,5 мкм и ширину около 2,5-3,5 мкм.
Нормальное соотношение длины и ширины составляет 1,5-1,7. При микроскопии в светлом поле наиболее часто
выявляемые аномалии головки — дефекты ее формы и размеров, включая увеличение, уменьшение, заострение
головки, грушевидную форму головки, аморфную головку, вакуолизацию головки (когда более 20% ее
поверхности занимают неокрашиваемые вакуоли), двойную головку, а также любые комбинации перечисленных
дефектов. Акросома сперматозоидов.
Рис. Ультраструктура нормальных сперматозоидов
в сканирующем (а) и трансмиссионном (б)
электронном микроскопе. * 10 000.
Г - головка;
Ш - шейка;
А - акросома;
Я - ядро с конденсированным хроматином;
Ц - цитоплазматическая капля.
6.
Слайд 5Строение головки сперматозоида
На ультрамикроскопическом уровне в головке сперматозоида различимы
акросомальная и
постакросомальная зоны.
Большую часть головки занимает ядро, заполненное конденсированным хроматином.
Пространство между ядром и передним
участком плазмолеммы спермия занято
акросомальной вакуолью — специализированной
лизосомой, содержащей группу
ферментов-лизинов оболочек яйцеклетки:
-акрозин (разрушает прозрачную зону овоцита),
-пенетраза (диссоциирует клетки лучистого венца),
гиалуронидаза (расщепляет гиалуроновую
кислоту), кислая фосфатаза (разрушает
форсхолин при прохождении спермиев через
плазмолемму овоцита).
7.
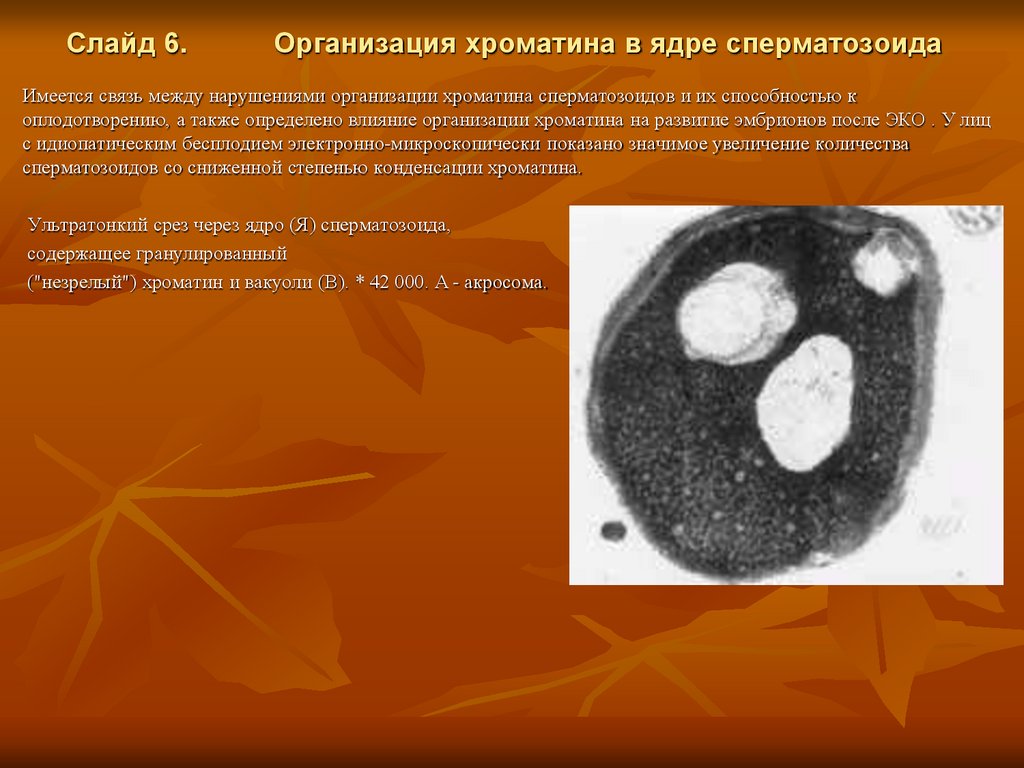
Слайд 6.Организация хроматина в ядре сперматозоида
Имеется связь между нарушениями организации хроматина сперматозоидов и их способностью к
оплодотворению, а также определено влияние организации хроматина на развитие эмбрионов после ЭКО . У лиц
с идиопатическим бесплодием электронно-микроскопически показано значимое увеличение количества
сперматозоидов со сниженной степенью конденсации хроматина.
Ультратонкий срез через ядро (Я) сперматозоида,
содержащее гранулированный
("незрелый") хроматин и вакуоли (В). * 42 000. А - акросома.
8.
Слайд 7Акросома
Акросома представлена комплексом Гольджи и покрывает около 2/3 передней поверхности головки.
Апикальное уплотнение, имеющееся у многих других видов, у человека отсутствует, однако видно
равномерное утолщение/истончение к экваториальному сегменту, и в целом она покрывает 40-70% головки
сперматозоида. При визуализации с помощью сканирующего электронного микроскопа видна окружающая
головку (т.е. акросомальный и постакросомальный регионы) бороздка, которая делит ее на две неравные части.
Экваториальный сегмент, за которым следует постакросомальный регион, визуализируется при этом нечетко.
Максимальной толщины и ширины сперматозоид достигает именно в постакросомальном регионе.
Акросома выглядит как шапочкообразная структура, покрывающая передние 2/3 головки сперматозоида, и
происходит из комплекса Гольджи сперматиды по мере ее дифференцировки в сперматозоид. В акросоме
содержится несколько гидролитических ферментов, в том числе гиалуронидаза и проакрозин, необходимых
для оплодотворения.
Задние отделы головки покрыты однослойной мембраной, называемой постнуклеарной шапочкой. Область
перекрытия акросомы и постнуклеарной шапочки образует экваториальный сегмент, который не участвует в
акросомальной реакции. Ядро, составляющее 65% всей головки, состоит из ДНК, ассоциированной с белками.
Хроматин плотно упакован, отдельные хромосомы неразличимы. Генетическая информация, в том числе
определяющие пол хромосомы X и Y, закодирована и хранится в виде ДНК.
При формировании акросомы несколько проакросомных гранул сливаются в одну большую гранулу, которая
контактирует с ядерной оболочкой. Она распластывается на поверхности ядра и формирует "шапочку",
занимающую 25-60% ядерной поверхности. В этот период гликопротеины переносятся из комплекса Гольджи
в акросому посредством мембранных пузырьков.
9.
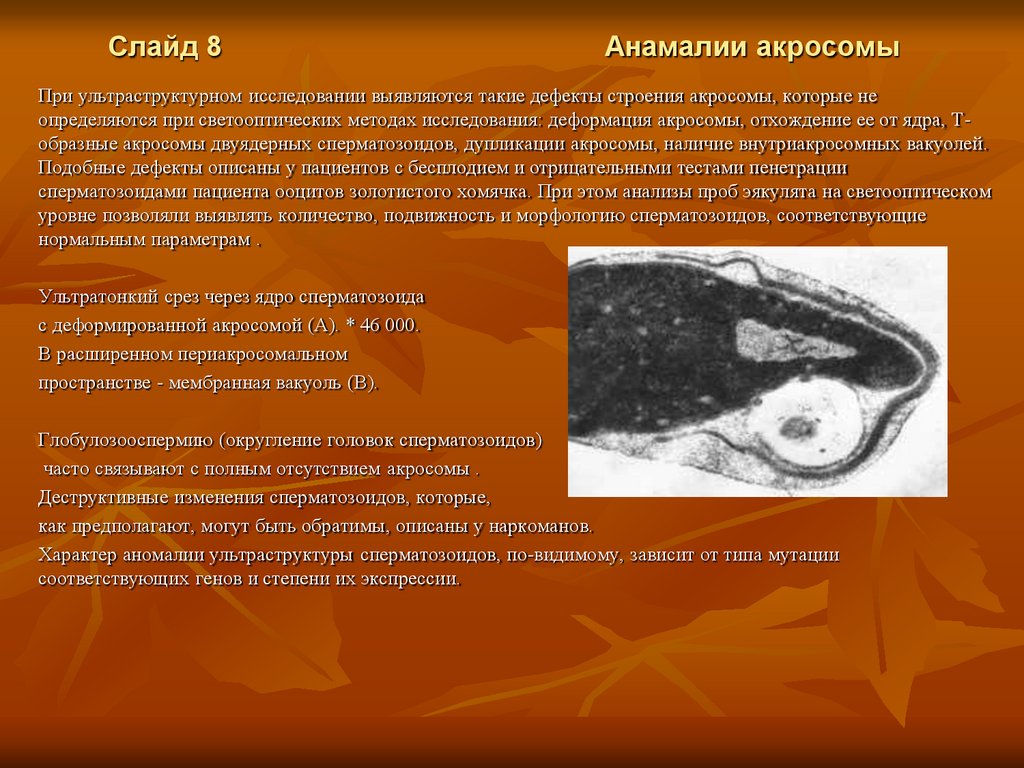
Слайд 8Анамалии акросомы
При ультраструктурном исследовании выявляются такие дефекты строения акросомы, которые не
определяются при светооптических методах исследования: деформация акросомы, отхождение ее от ядра, Тобразные акросомы двуядерных сперматозоидов, дупликации акросомы, наличие внутриакросомных вакуолей.
Подобные дефекты описаны у пациентов с бесплодием и отрицательными тестами пенетрации
сперматозоидами пациента ооцитов золотистого хомячка. При этом анализы проб эякулята на светооптическом
уровне позволяли выявлять количество, подвижность и морфологию сперматозоидов, соответствующие
нормальным параметрам .
Ультратонкий срез через ядро сперматозоида
с деформированной акросомой (А). * 46 000.
В расширенном периакросомальном
пространстве - мембранная вакуоль (В).
Глобулозооспермию (округление головок сперматозоидов)
часто связывают с полным отсутствием акросомы .
Деструктивные изменения сперматозоидов, которые,
как предполагают, могут быть обратимы, описаны у наркоманов.
Характер аномалии ультраструктуры сперматозоидов, по-видимому, зависит от типа мутации
соответствующих генов и степени их экспрессии.
10.
Слайд 9Шейка сперматозоидов.
Шейка сперматозоидов. Шейка соединяет головку и хвост. Она очень ломкая, поэтому наличие в эякуляте
декапитированных сперматозоидов встречают довольно часто.
Шейка сперматозоида имеет сложное строение. На тотальном препарате она выявляется как утолщение
жгутика. У сформированного сперматозоида шейка представляет собой усеченный конус, содержащий
проксимальную и измененную дистальную центриоли. Усеченный конус шейки на уровне дистальной центриоли
состоит из 9 удлиненных поперечно исчерченных столбиков. Эти столбики напротив центриолярного отростка
сливаются, теряют индивидуальные очертания и совместно продолжаются по направлению к головке, сливаясь с
базальной пластинкой, лежащей у основания имплантационной ямки.
Рис. Схематическое изображение шейки сперматозоида (а)
и ультратонкий срез (б) через шейку нормального
сперматозоида. * 13 000.
М - митохондрии.
Промежуточный отдел содержит аксонему и 9 продольно ориентированных
элементов цитоскелета, состоящих из кератиноподобных белков и являющихся
продолжением сегментированных колонн. Кнаружи от этих волокон располагаются
митохондрии. Аксонема построена по принципу реснички, биения которой
инициируются катионами кальция, находящимися в окружающей среде, и
митохондриальной АТФ спермия. В концевом отделе хвоста элементы цитоскелета исчезают, и аксонема
прикрыта лишь плазмолеммой. У человека дуплеты аксонемы в концевом отделе распадаются на 18 одиночных
микротрубочек.
Центриоли человека, наследуются через отцовские половые клетки. Функциональный центр образования
микротрубочек астросферы спермия человека связан с центриолями сперматозоидов. Так, у диспермических
зигот формируются трехполюсные веретена дробления. A. Sathananthan высказал гипотезу о том, что если
дефектные центриоли сперматозоидов наследуются эмбрионом при оплодотворении, они могут приводить к
аномальному дроблению и развитию аномальных эмбрионов. Нет прямых доказательств этой гипотезы у
человека.
11.
Слайд10Аномалии шейки сперматозоида
Аномалией строения шейки является наличие остаточных цитоплазматических капель,
размер которых превышает величину головки. К патологическим изменениям шейки
сперматозоидов относят гетероаксиальность - те случаи, когда головка сперматозоида
и жгутик расположены под углом друг к другу.
Рис. Сперматозоид с цитоплазматической каплей на шейке
Рис. Сперматозоид с гетероаксиальностью (отклонение головки (Г) от оси
симметрии менее 90°) в трансмиссионном (б; * 35 000) электронном
микроскопе.
Рис. Сперматозоид с гетероаксиальностью (отклонение головки (Г) от оси
симметрии более 90 трансмиссионном (б; Ѕ 36 000) электронном микроскопе.
В зоне шейки наблюдается проксимальная центриоль (Ц),
остальные структуры соединительного участка отсутствуют.
12.
Слайд11Концевой отдел сперматозоидов
Концевой отдел сперматозоидов. Хвост появляется на стадии сперматид Концевой отдел плохо различим при
световой микроскопии. Оболочка и грубоволокнистые филаменты отсутствуют. Хвост, содержащий
двигательный аппарат, имеет длину 40-50 мкм и берет начало от центриоли сперматиды. Его волнообразные
движения генерируются в районе шейки и распространяются в дистальном направлении по типу хлыстового
удара. При световой микроскопии чаще всего выявляют следующие отклонения в строении шейки и среднего
отдела: их отсутствие, искривление хвоста, удлиненный средний отдел, неправильная форма/искривление
среднего отдела, истончение среднего отдела (отсутствие митохондриальной оболочки), а также различные
сочетания указанных дефектов. Аномалии хвоста включают укорочение, удвоение/утроение, деформацию в виде
шпильки, разрывы, неравномерную ширину, патологическую извитость с терминальными капельками, а также
различные сочетания указанных дефектов. Цитоплазматические капли, занимающие более 1/3 объема головки
сперматозоида, считают отклонением от нормы. Они обычно располагаются в области шейки/среднего отдела
хвоста, хотя у некоторых незрелых сперматозоидов могут находиться и в других участках хвоста.
По данным сканирующей электронной микроскопии,
в хвосте выделяют три отдела: средний, основной и
концевой. В среднем отделе четко различимы
митохондриальные спирали. Они резко обрываются
в начале среднего отдела. К заднему концу средний
отдел сужается. Видны продольный стержень и
поперечные ребра. Короткий концевой отдел имеет
меньший диаметр из-за отсутствия наружных волокон.
При трансмиссионной электронной микроскопии видно,
что в среднем отделе существует цитоплазматическая часть
и богатая липидами митохондриальная оболочка, состоящая
из нескольких спиральных митохондрий, выстроенных вокруг аксиального филамента в форме «завитков
улитки». Средний отдел обеспечивает сперматозоид энергией, необходимой для движения. Дополнительное
наружное кольцо из 9 грубоволокнистых фибрилл окружает центральный стержень, состоящий из 11 фибрилл.
13.
Слайд 12Хвостовой отдел сперматозоидов
Морфологическую основу жгутика составляет аксонема, состоящая из микротрубочек, организованных по
универсальной схеме 9+2, характерной для большинства типов ресничек и жгутиков клеток эукариотических
организмов. Аксонема тянется вдоль всей длины жгутика. В среднем отделе аксонема жгутика окружена слоем
из 9 дополнительных фибрилл (9+9+2), кнаружи от которых расположены митохондрии. В основном отделе они
заменяются фиброзным слоем, в концевом отделе остается только аксонема.
Схематическое изображение аксонемы жгутика нормального сперматозоида на поперечном срезе (а).
Ультратонкие поперечные
срезы через основной
участок жгутика пациента
с нормозооспермией (б)
и сперматозоида пациента Б
с астенозооспермией (в). * 145 000.
В аксонеме жгутика сперматозоида
на рис. 6, в полностью отсутствуют динеиновые ручки периферических дуплетов микротрубочек.
Девять грубоволокнистых фибрилл наружного кольца уменьшаются в толщине и в конце концов исчезают,
оставляя только внутренние фибриллы на аксиальном стержне на протяжении почти всей длины основного
отдела. Фибриллы основного отдела окружены фиброзной оболочкой хвоста, состоящей из разветвленных и
анастомозирующих между собой полукруглых тяжей, или ребер, удерживаемых вместе за счет прикрепления к
двум полоскам, тянущимся по обеим сторонам хвоста на всем его протяжении. Хвост заканчивается концевым
отделом длиной 4-10 мкм и диаметром менее 1 мкм. Столь малый диаметр обусловлен отсутствием наружной
фиброзной оболочки и дисталь-ным истончением микротрубочек.
14.
Слайд 13Анамалии хвостового отдела спериатозоидов
Наиболее частым дефектом строения аксонемы является отсутствие одной или обеих центральных
микротрубочек (дефект 9 + 1 или 9 + 0), что выражается в значительном уменьшении или полном отсутствии
подвижности сперматозоидов. Достаточно часто встречаются такие аномалии аксонемы, как:
1) отсутствие динеиновых ручек около периферических дуплетов микротрубочек (и наружных и внутренних,
либо только наружных или только внутренних);
2) дезорганизация какого-либо периферического дуплета;
3) отсутствие радиальных спиц аксонемы
Сперматозоиды с дефектами строения аксонемы не способны проходить через женские половые пути и
достигать верхних отделов маточных труб, однако они дают акросомную реакцию и пенетрируют ооциты.
Структурные аномалии дополнительных фибрилл встречаются реже, чем аномалии аксонемы, и могут
выражаться в изменении их формы хвостового отдела.
15.
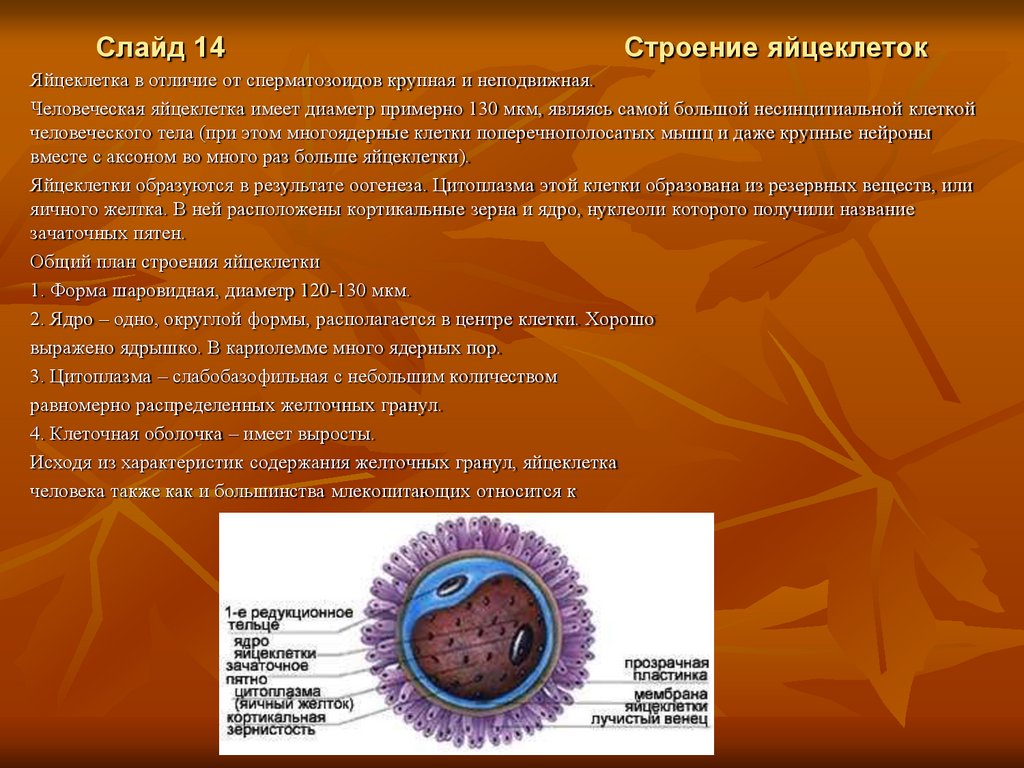
Слайд 14Строение яйцеклеток
Яйцеклетка в отличие от сперматозоидов крупная и неподвижная.
Человеческая яйцеклетка имеет диаметр примерно 130 мкм, являясь самой большой несинцитиальной клеткой
человеческого тела (при этом многоядерные клетки поперечнополосатых мышц и даже крупные нейроны
вместе с аксоном во много раз больше яйцеклетки).
Яйцеклетки образуются в результате оогенеза. Цитоплазма этой клетки образована из резервных веществ, или
яичного желтка. В ней расположены кортикальные зерна и ядро, нуклеоли которого получили название
зачаточных пятен.
Общий план строения яйцеклетки
1. Форма шаровидная, диаметр 120-130 мкм.
2. Ядро – одно, округлой формы, располагается в центре клетки. Хорошо
выражено ядрышко. В кариолемме много ядерных пор.
3. Цитоплазма – слабобазофильная с небольшим количеством
равномерно распределенных желточных гранул.
4. Клеточная оболочка – имеет выросты.
Исходя из характеристик содержания желточных гранул, яйцеклетка
человека также как и большинства млекопитающих относится к
16.
Слайд 15Структурно-функциональные особенности яйцеклетки
1. Ядро содержит гаплоидный набор хромосом.
2. Цитоплазма характеризуется региональностью биогенетических свойств (овотипическая детерминация).
Вероятно на этом уровне детерминации формируется и презумптивный (praesumptio - предназначение,
предположение) субстрат «зародышевой плазмы» – источника образования
гонобластов.
3. В цитоплазме имеется несколько видов гранул (мембранных пузырьков), компоновка которых происходит в
комплексе Гольджи:
а) желточные гранулы – заполнены белками вителлинами (vitellus желток), которые являются основным
трофическим материалом яйцеклетки,
б) кортикальные гранулы – заполнены гликозаминогликанами (ГАГсложные углеводы), находятся на
периферии цитоплазмы в подмембранном слое,
в) мультивезикулярные (лизосомоподобные) тельца – заполнены протеолитическими ферментами, синтез
которых происходит в ЭПС. Кортикальные гранулы и мультивезикулярные тельца при внедрении
сперматозоида подвергаются экзоцитозу (кортикальная реакция).
Вследствие этого формируется специальная оболочка оплодотворения. Она предотвращает полиспермию
(многоспермное оплодотворение).
4. Хорошо развиты многие органеллы общего назначения:
а) митохондрии (до 400-500 шт.) общесоматического типа, наследуются зиготой и в дальнейшем клетками всего
зарождающегося организма,
б) комплекс Гольджи – участвует в гранулообразовании,
в) ЭПС (шероховатая и гладкая) – обеспечивает внутриклеточные синтезы,
г) свободные рибосомы – белковые синтезы внутреннего потребления
5. Имеются мембранные рецепторы (рецепторы фертильности) индивидуального и видового отбора
сперматозоидов. Представлены специализированными белками в составе плазмолеммы. Отбор
осуществляется по принципу комплементарности с соответствующим рецептором сперматозоида.
17.
Слайд 166. Обладает структурным фактором иммунной защиты. Представлен блестящей оболочкой (модифицированный
гликокаликс), в которой имеются каналы для отростков фолликулярных клеток (см. ниже).
7. Жизнеспособность яйцеклетки после овуляции (выхода гаметы из яичника) весьма ограничена. Способность
к оплодотворению сохраняется в течение суток в оптимальных средах обитания. Таковыми являются: серозная
жидкость ампул маточных труб и перитонеальная жидкость
прямокишечно-маточного (Дугласова) пространства, а также искусственные субстраты, используемые для
экстракорпоральных оплодотворений
На ультрамикроскопическом уровне яйцеклетка человека имеет крупное округлое ядро, в котором преобладает
эухроматин, и цитоплазму с умеренным количеством митохондрий с немногочисленными ламеллярными
кристами. Хорошо развита система белкового синтеза из-за высокого содержания рибо- и полисом, коротких
канальцев гранулярной эндоплазматической сети. В цитоплазме редко и диффузно расположены
немногочисленные скопления вителлиновых гранул — коллекторов трофических включений.
У высших плацентарных млекопитающих и человека женские половые клетки не существуют изолированно. За
исключением самых ранних стадий развития (первичных половых клеток, овогоний) они находятся в тесном
контакте с соматическими клетками яичника (фолликулярными эпителиоцитами и соединительнотканными
клетками), которые формируют вокруг каждой половой клетки эпителиальную и соединительнотканную
оболочки. Комплекс "половая клетка — соматические оболочки" именуется овариальным фолликулом, или овосоматическим (фолликулярным) гистионом. Строение овариального фолликула усложняется в процессе
овогенеза.
18.
Слайд 17Оболочки яйцеклетки
Наружная оболочка клетки имеет многослойную структуру. Каждая из составляющих выполняет свои основные
функции. Оболочки устроены так, что после проникновения в клетку одного сперматозоида они становятся
совершенно непроницаемы для других мужских клеток. Одновременно с этим оболочки имеют достаточный запас
прочности, чтобы при транспортировке яйцеклетки по фаллопиевой трубе в полость матки после оплодотворения
дробящаяся клетка не пострадала и смогла благополучно завершить свое «путешествие» и имплантироваться.
1. Фолликулярная оболочка (наружная). Состоит из нескольких рядов фолликулоцитов – клеток эпителия
фолликулов яичника. Фолликулы – это специальные структуры яичника, в которых происходит развитие половых
женских клеток.
Функции оболочки:
а) трофическая (по отношению к яйцеклетке),
б)защитная; в) эндокринная; г) участие в синтезе компонентов блестящей оболочки.
2. Блестящая оболочка (прозрачная зона) – модифицированный гликокаликс. Расположена под фолликулярной
оболочкой. Zona pellucida, блестящая оболочка, представляет собой специфическую внеклеточного оболочку ооцита,
присущую млекопитающим. Толщина блестящей оболочки может изменяться от 1 до около 25 мкм в зависимости от
вида животного. Zona pellucida состоит из гликопротеинов (ZP1-3 , имеется дополнительный ZP4), которые
формируют сеть микрофибрилл, плотно упакованных у мембраны ооцита (оолемы) и более разреженно — на
периферии.
3. Лучистый венец технически не является производным яйцеклетки, а состоит из отдельных клеток, которые
называются кучевых (лат. Cumulus oophorus). Они окружают ооцит как в фолликуле, так и после овуляции. До
овуляции кучевые клетки является ближайшим к ооцита слоем гранулёзных клеток, функция которых заключается в
питании ооцита через специальные образования, трансзональни проекции, которые соединяют эти клетки.
4. Оолемма. Вителинова оболочка, или первичная оболочка состоит из белков, которые помогают сперматозоида
попасть к яйцеклетке и выполняют Видоспецифические барьерную функцию, обеспечивая попадание сперматозоида
только соответствующего вида. Вителинова оболочка присуща беспозвоночным и позвоночным, однако у
млекопитающих она специфическая и называется zona pellucida. Как у млекопитающих, так и у других организмов в
белков вителиновои оболочки (zona pellucida) имеющийся домен ZP, с помощью которого происходит
полимеризация белков и уплотнения оболочки.4. Оболочка оплодотворения - образуется только после
проникновения оплодотворяющего сперматозоида. Представляет собой утолщенную уплотнившуюся блестящую
19.
Слайд 18Функции яйцеклетки
Основной комплексной функцией яйцеклетки является функция оплодотворения. Она складывается из целого
ряда производных функции:
1. Секреторная – в состоянии активной жизнедеятельности яйцеклетка синтезирует и выделяет
низкомолекулярные биологические соединения – гиногамоны. Они определяют положительный хемотаксис
сперматозоида.
2. Осуществление индивидуального и видового отбора сперматозоидов.
3. Предотвращение полиспермного оплодотворения.
4. Обеспечение процессов овотипической детерминации ( презумпция «зародышевой плазмы» и
соматоплазмы).
5. Обеспечение процесса наследования зиготой митохондрий.
20.
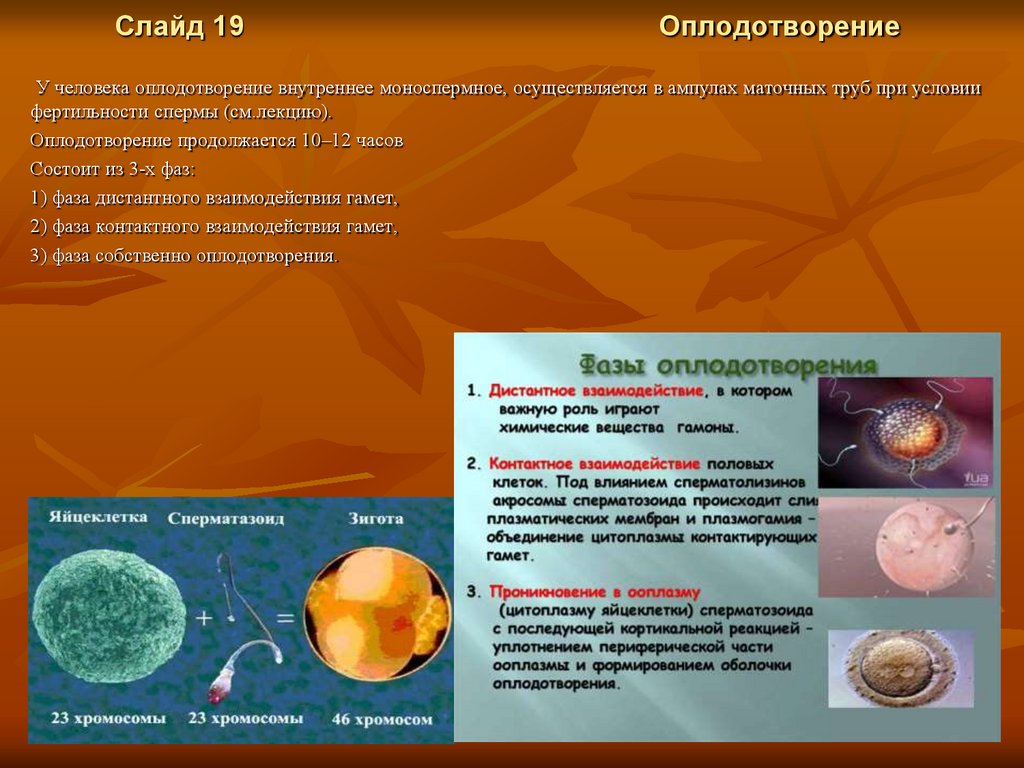
Слайд 19Оплодотворение
У человека оплодотворение внутреннее моноспермное, осуществляется в ампулах маточных труб при условии
фертильности спермы (см.лекцию).
Оплодотворение продолжается 10–12 часов
Состоит из 3-х фаз:
1) фаза дистантного взаимодействия гамет,
2) фаза контактного взаимодействия гамет,
3) фаза собственно оплодотворения.
21.
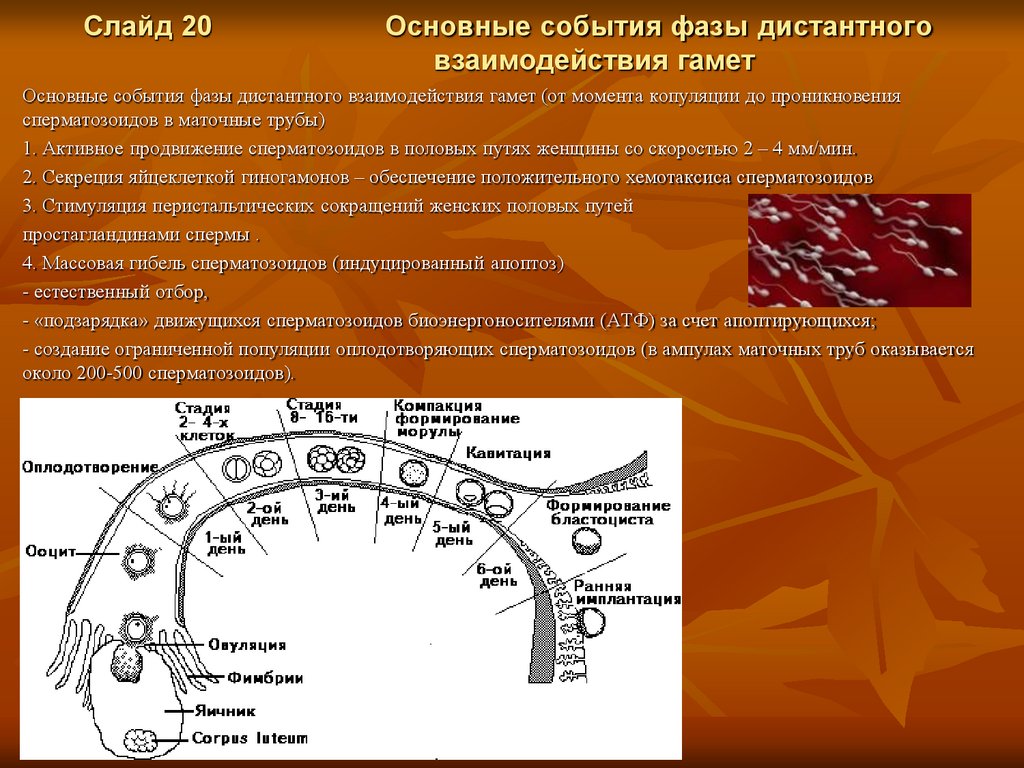
Слайд 20Основные события фазы дистантного
взаимодействия гамет
Основные события фазы дистантного взаимодействия гамет (от момента копуляции до проникновения
сперматозоидов в маточные трубы)
1. Активное продвижение сперматозоидов в половых путях женщины со скоростью 2 – 4 мм/мин.
2. Секреция яйцеклеткой гиногамонов – обеспечение положительного хемотаксиса сперматозоидов
3. Стимуляция перистальтических сокращений женских половых путей
простагландинами спермы .
4. Массовая гибель сперматозоидов (индуцированный апоптоз)
- естественный отбор,
- «подзарядка» движущихся сперматозоидов биоэнергоносителями (АТФ) за счет апоптирующихся;
- создание ограниченной популяции оплодотворяющих сперматозоидов (в ампулах маточных труб оказывается
около 200-500 сперматозоидов).
22.
Слайд 21Основные события фазы контактного взаимодействия
гамет
(происходят в ампулах маточных труб)
1. Пассивное вращение женской половой клетки (овоцит второго порядка) под действием
перистальтики маточных труб и циркуляции в них серозной жидкости, удаление фолликулярной
оболочки, обнажение рецепторов фертильности.
2. Капацитация сперматозоидов – удаление гликокаликса, обнажение рецептора фертильности.
3. Взаимодействие рецепторов фертильности по принципу комплементарности; индивидуальный
отбор сперматозоида.
4. Акросомальная реакция и пенетрация (проникновение).
5. Внедрение головки сперматозоида в цитоплазму овоцита.
6. Кортикальная реакция и образование оболочки оплодотворения.
23.
Слайд 22Акросомная реакция
Акросомная реакция — разновидность экзоцитоза. При акросомной реакции наружная мембрана акросомы и
клеточная мембрана сливаются и формируют мелкие пузырьки, отделяющиеся от головки сперматозоида.
При этом из акросомы освобождаются гиалуронидазы, протеазы (в том числе акрозин), гликозидазы, липазы,
нейраминидаза и фосфатазы. Ферменты расщепляют молекулы блестящей оболочки, что позволяет
сперматозоиду преодолеть этот барьер.
Корткаьная реакция протекает в несколько этапов.
1.Контакт со студенистой оболочкой яйцеклетки.
2.Слившиеся мембраны разрываются, образуя отверстия, через которые изливается содержимое акросомы;
ферменты акросомы разрушают яйцеклетку, открывая доступ к желточной оболочке.
3.Внутренний участок акросомы начинает быстро выпячиваться за счёт образования активных филаментов;
формируется акросомный вырост.
4.Акросомный вырост вступает в контакт с поверхностью желточной оболочки, при этом происходит
комплементарное взаимодействие
белка бендина, который встроен в
мембрану акросомного выроста и
рецепторов в желточной оболочке.
5.Вслед за реакцией узнавания и реакцией
фермент-рецептор желточная оболочка
разрушается и на плазмолемме яйцеклетки
образуется вырост, направленный
навстречу акросомной нити – бугорок
оплодотворения.
24.
Слайд 23Основные события фазы собственно
оплодотворения
Основные события фазы собственно оплодотворения (начинается после внедрения головки сперматозоида в
цитоплазму овоцита второго порядка и развития кортикальной реакции).
1. Внесение ядра сперматозоида (мужского пронуклеуса) и его центриолей в овоплазму. В мужском и женском
пронуклеусах содержится гаплоидный набор хромосом.
2. Овоцит превращается в яйцеклетку (завершается фаза созревания овогенеза)
3. Формирование полярности яйцеклетки по вектору проникновения сперматозоида. Образуется
краниокаудальная ось будущего организма (овотипический уровень детерминации).
4. Сближение ( 12 ч.) пронуклеусов.
5. Слияние (синкарион) пронуклеусов. Образование единой кариолеммы
и диплоидного набора хромосом.
6. Депрессирование (разблокирование) гена пролиферации.
7. Формирование зиготы, которая совершает первое деление дробления
25.
Слайд 24Проникновение сперматозоида в яйцеклетку
Для человека характерна моноспермия, т.е.проникает только один сперматозоид. Сразу в яйцеклетке
происходит кортикальная реакция: под цитолеммой лопаются кортикальные гранулы, из них освобождаются
гликопротеиды, которые образуют оболочку оплодотворения. Она отличается по составу от блестящей
оболочки и препятствует полиспермии, т. к. сперматозоиды не могут ее растворить.
Слияние ядер гамет проходит в 4 стадии:
1 – стадия 2-х пронуклеусов (в цитоплазму яйцеклетки проникают ядро и центриоли сперматозоида), 2 – стадия
синкариона (двойное ядро),
3 – стадия слияния ядер в одно диплоидное ядро (образуется зигота),
4-я стадия – ооплазматическая сегрегация (в цитоплазме зиготы формируются презумптивные зачатки).
Затем зигота приступает к дроблению, во время которого зародыш продвигается к матке. Д
26.
Слайд 25Дробление
Дробление - череда последовательных митотических делений зиготы, в результате которого образуются
эмбриональные клетки – бластомеры (blastos- зачаток, meros- часть). Дробление – это специальная
разновидность митоза с практическим отсутствием пресинтетического периода.
Бластомеры (в отличии от соматических клеток) обладают рядом свойств:
1) не расходятся;
2) не растут;
3) не дифференцируются;
4) Не функционируют;
5) не апоптируют.
Дробление у человека происходит в процессе продвижения эмбриона по маточной трубе. Оно длится
приблизительно 5-6 суток. В итоге дробления формируется многоклеточный зародыш – бластоциста.
Дробление начинается к концу 1-х суток и проходит под оболочкой оплодотворения. У человека дробление
полное, неравномерное. Вначале образуются 2 бластомера – темный и светлый. Темные крупные, дробятся
медленнее, из них образуется эмбриобласт – материал зародыша и внезародышевых органов. Светлые – мельче,
дробятся быстрее, обрастают вокруг темных и дадут начало трофобласту. До 4 суток дробление
идет медленно, 1 деление в сутки. Образуется 2, 3, 5, 7 бластомеров. К 4 суткам зародыш состоит из 8–14
бластомеров. Первые 8 бластомеров полипотентны, и каждый может дать начало целому организму. Затем
дробление идет быстро, и на 5 сутки зародыш состоит из 100–107 бластомеров. Вначале бластомеры
образуют плотный узелок, похожий на морулу, но он устроен сложнее и называется стерробластула (бластула
без полости). Внутри узелка содержатся темные бластомеры эмбриобласта, а снаружи – один слой светлых
бластомеров трофобласта. Трофобласт всасывает жидкость из секрета яйцеводов и выделяет ее внутрь
зародыша. Зародыш становится в виде пузырька и называется бластоциста. В период дробления зародыш
плавает в секрете яйцевода и за счет перистальтики продвигается к матке, на 5–6 сутки в виде бластоцисты он в
матку и до 7 суток плавает в секрете маточных желез. Этот период называется стадией свободной бластоцисты.
27.
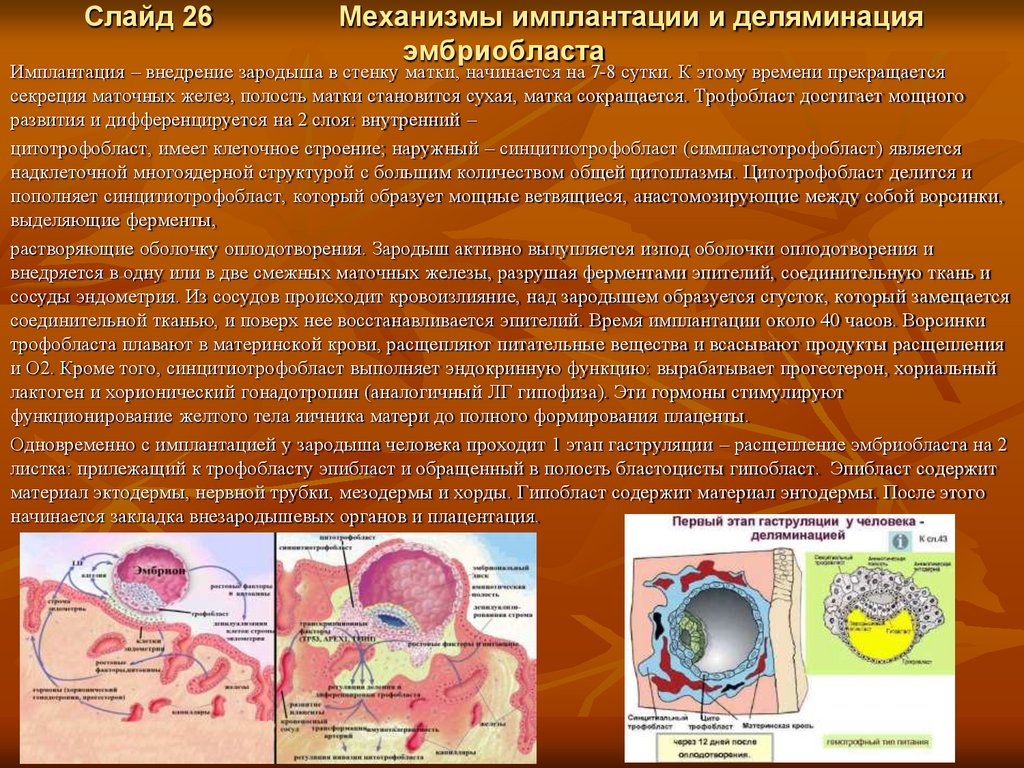
Слайд 26Механизмы имплантации и деляминация
эмбриобласта
Имплантация – внедрение зародыша в стенку матки, начинается на 7-8 сутки. К этому времени прекращается
секреция маточных желез, полость матки становится сухая, матка сокращается. Трофобласт достигает мощного
развития и дифференцируется на 2 слоя: внутренний –
цитотрофобласт, имеет клеточное строение; наружный – синцитиотрофобласт (симпластотрофобласт) является
надклеточной многоядерной структурой с большим количеством общей цитоплазмы. Цитотрофобласт делится и
пополняет синцитиотрофобласт, который образует мощные ветвящиеся, анастомозирующие между собой ворсинки,
выделяющие ферменты,
растворяющие оболочку оплодотворения. Зародыш активно вылупляется изпод оболочки оплодотворения и
внедряется в одну или в две смежных маточных железы, разрушая ферментами эпителий, соединительную ткань и
сосуды эндометрия. Из сосудов происходит кровоизлияние, над зародышем образуется сгусток, который замещается
соединительной тканью, и поверх нее восстанавливается эпителий. Время имплантации около 40 часов. Ворсинки
трофобласта плавают в материнской крови, расщепляют питательные вещества и всасывают продукты расщепления
и О2. Кроме того, синцитиотрофобласт выполняет эндокринную функцию: вырабатывает прогестерон, хориальный
лактоген и хорионический гонадотропин (аналогичный ЛГ гипофиза). Эти гормоны стимулируют
функционирование желтого тела яичника матери до полного формирования плаценты.
Одновременно с имплантацией у зародыша человека проходит 1 этап гаструляции – расщепление эмбриобласта на 2
листка: прилежащий к трофобласту эпибласт и обращенный в полость бластоцисты гипобласт. Эпибласт содержит
материал эктодермы, нервной трубки, мезодермы и хорды. Гипобласт содержит материал энтодермы. После этого
начинается закладка внезародышевых органов и плацентация.
28.
Контрольные вопросы1.Строение головки сперматозоида.
2..Строение шейки сперматозоида.
3.Строение хвостика сперматозоида.
4. Строение акросомы, ее патологии.
5. Строение оболочек яйцеклетки.
6. Структурно-функционаьные особенности яйцеклетки.
7. Основные события дистанционного взаимодействия гамет.
8. Основные события контактного взаимодействия гамет.
9. Опищите фазы акросомальной реакции.
10. Основные события процесса оплодотворения.
11. Особенности процесса оплодотворения.