



























 biology
biologySimilar presentations:




")





Активность скелетных мышц в условиях естественной деятельности
1. ОФВТ-2
Берг М.Д.ОФВТ-2
Активность скелетных мышц в
условиях естественной деятельности
2.
План лекцииАктивность скелетных мышц в условиях естественной
деятельности
1. Функции скелетных мышц.
2. Двигательные (моторные) единицы.
3. Виды сокращений скелетных мышц, оценка сократимости.
4. Механизм сокращения и расслабления.
5. Сопряжение между возбуждением и сокращением.
6. Энергия для сокращения.
Физиология гладких мышц
7. Функции гладких мышц. Структурно-функциональная единица.
8. Особенности сокращения и расслабления. Источники кальция.
9. Виды сокращений. Автоматия гладких мышц.
10. Электрические явления в гладкой мышце.
Физиология секреторных клеток
11. Секреторные клетки, способы и виды секреции, механизм
секреции.
Профессор Берг М.Д.
3.
До 70% массы тела приходится на мышцы.Мышцы – это биологические машины, преобразующие
энергию АТФ в работу. КПД = 20-30%, может достигать
40-50%.
Виды мышц: скелетная, сердечная, гладкая.
Функции скелетных мышц:
1) Поддержание позы и равновесия.
2) Движение тела и его частей в пространстве бытовые,
рабочие, спортивные движения).
3) Коммуникации между людьми и животными (мимика,
жесты, речь).
4) Модулирование функционального состояния
систем организма, уровня здоровья, продолжительности жизни, психоэмоционального состояния,
иммунитета, нейрогенеза путем рефлекторных
(М.Р.Могендович) и гуморальных (B.K.Pedersen)
влияний.
Профессор Берг М.Д.
4.
Структурно-функциональная единица – двигательнаяединица (ДЕ) или моторная единица (МЕ).
Это двигательный нейрон и группа мышечных волокон,
которые он иннервирует. Количество мышечных
волокон в ДЕ различно.
Быстрые ДЕ имеют от нескольких единиц до нескольких
десятков мышечных волокон. Они обеспечивают
быстрые движения и коммуникации.
Медленные ДЕ имеют сотни и до 2-х тысяч мышечных
волокон. Они поддерживают позу и равновесие,
длительные усилия.
Каждая ДЕ объединяет свой тип мышечных волокон:
красные окислительные (I), белые гликолитические (IIb)
или смешанный окислительно-гликолитический тип (IIa).
Мышечные волокна, принадлежащие различным ДЕ,
рассредоточены по всей мышце.
Профессор Берг М.Д.
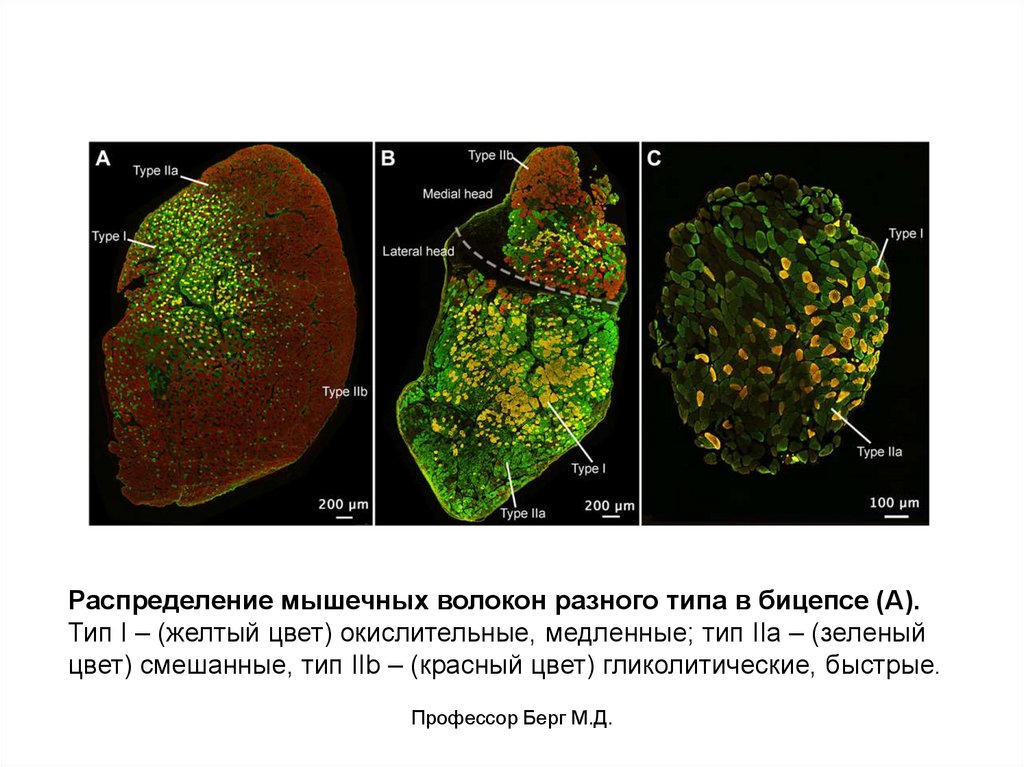
5.
Распределение мышечных волокон разного типа в бицепсе (А).Тип I – (желтый цвет) окислительные, медленные; тип IIa – (зеленый
цвет) смешанные, тип IIb – (красный цвет) гликолитические, быстрые.
Профессор Берг М.Д.
6.
Мотонейрон генерирует ПД с частотой 20-50 Гц,которые распространяются по его аксону. В месте
контакта с мышечным волокном образуется синапс,
через который возбуждение передается химическим
путем с помощью ацетилхолина и никотинового
холинорецептора (Харуки Мураками «Подземка»). Возникает
ПД мышцы, а затем её сокращение.
Особенности нервных влияний:
• всегда серия импульсов,
• одновременно ко всем мышечным волокнам данной
ДЕ,
• к разным ДЕ – не одновременно, в результате –
суммация сокращений мышечных волокон (гладкий
тетанус),
• для увеличения силы сокращений: > частоты ПД, >
числа ДЕ, вовлечение в работу ДЕ первого, второго и
третьего типов (спортивные
рекорды).
Профессор Берг
М.Д.
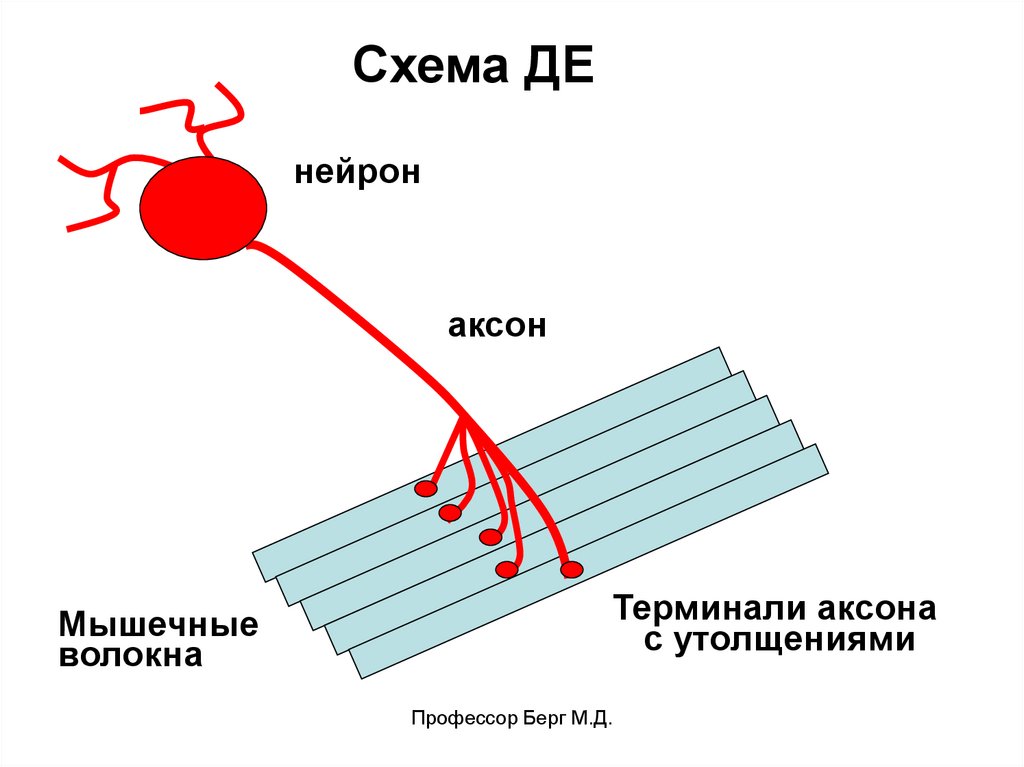
7.
Схема ДЕнейрон
аксон
Мышечные
волокна
Терминали аксона
с утолщениями
Профессор Берг М.Д.
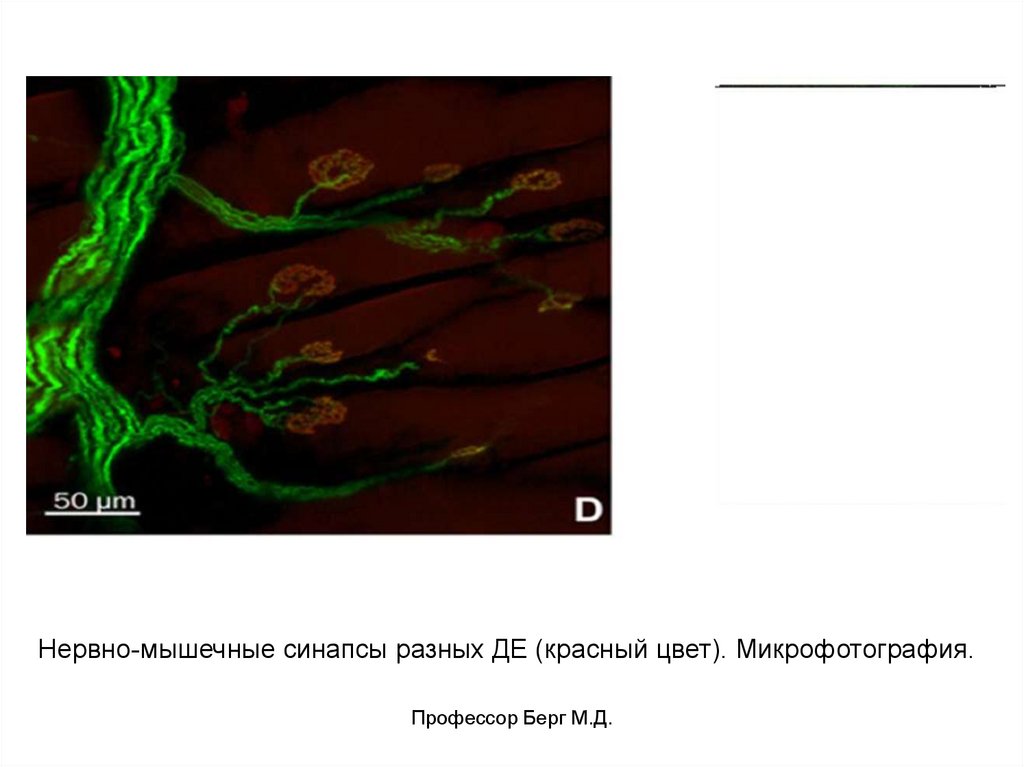
8.
Нервно-мышечные синапсы разных ДЕ (красный цвет). Микрофотография.Профессор Берг М.Д.
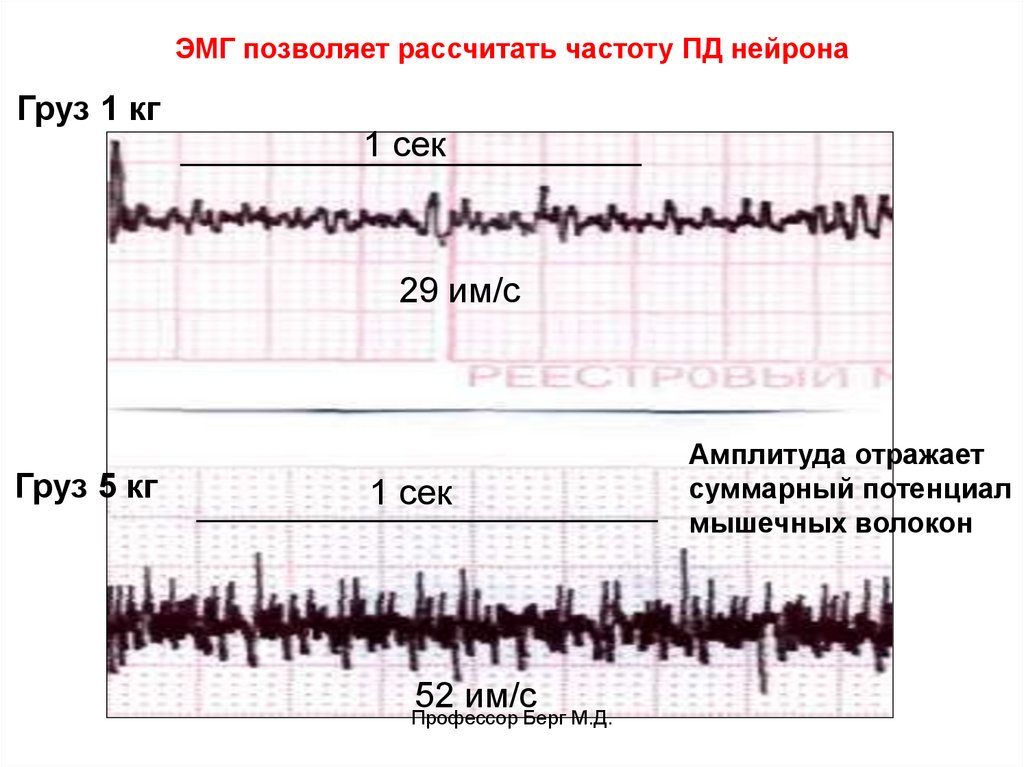
9.
ЭМГ позволяет рассчитать частоту ПД нейронаГруз 1 кг
1 сек
29 им/с
Груз 5 кг
1 сек
52 им/с
Профессор Берг М.Д.
Амплитуда отражает
суммарный потенциал
мышечных волокон
10.
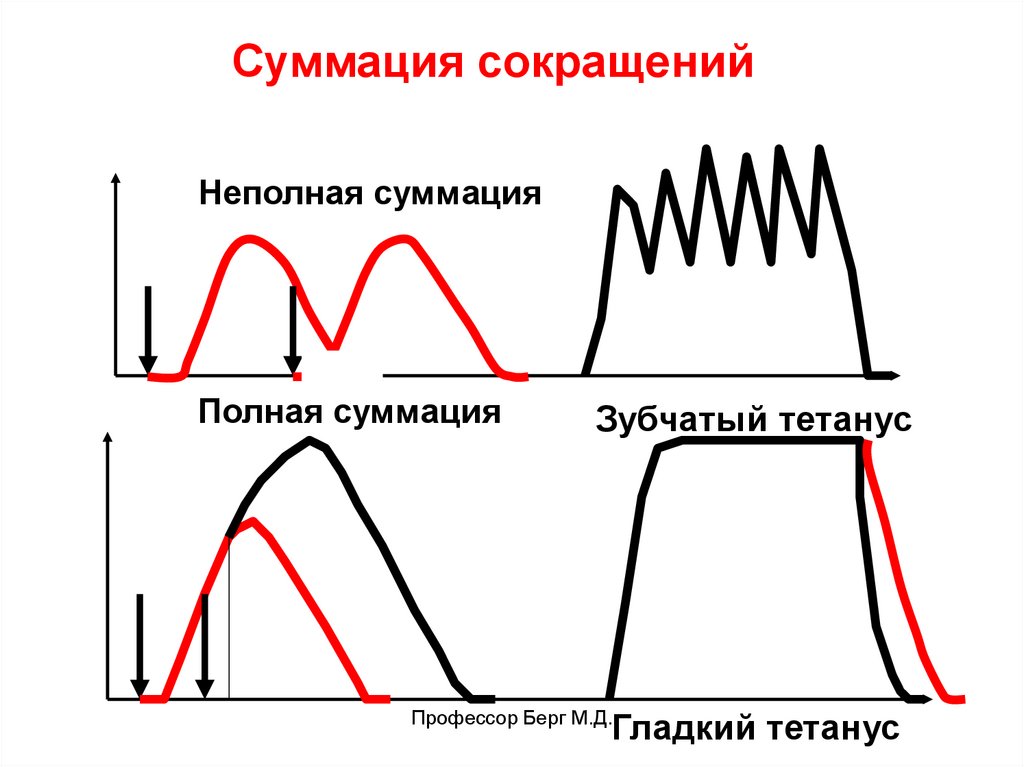
Виды сокращений скелетных мышц:1) Тонические (нейрогенный тонус медленных ДЕ),
2) Ритмические:
• одиночные,
• тетанические (зубчатый и гладкий тетанус),
в естественных условиях – гладкий тетанус.
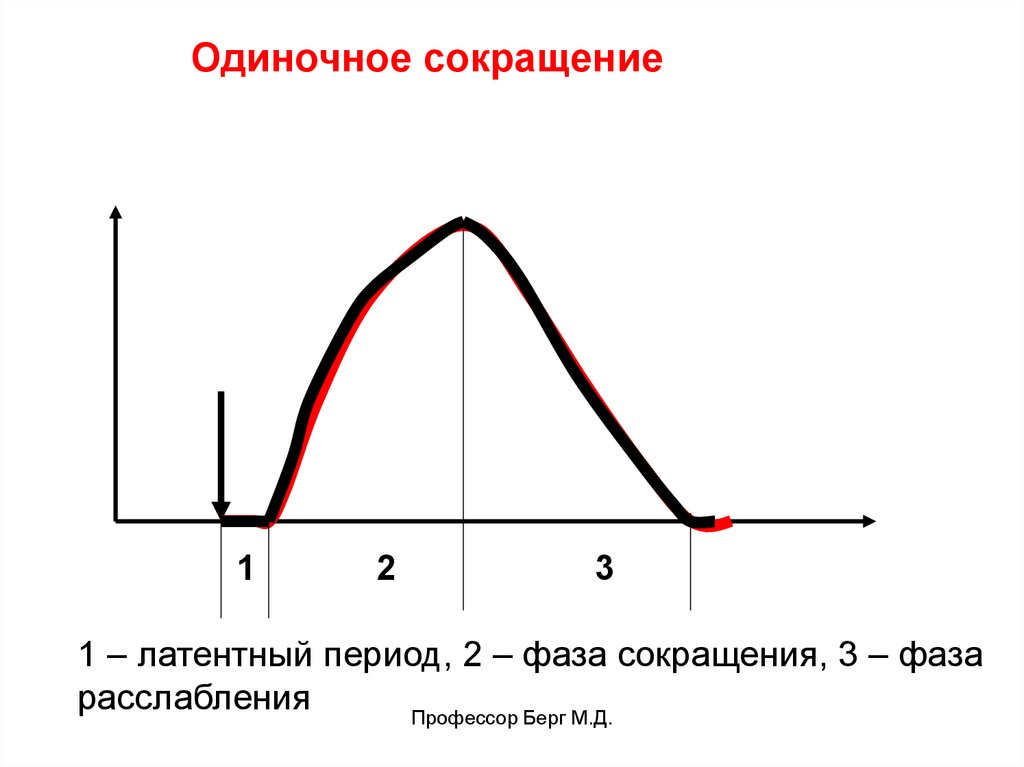
Одиночное сокращение
возникает на однократное раздражение (только в
эксперименте). Имеет фазы:
• латентный период,
• фазу сокращения,
• фазу расслабления.
Профессор Берг М.Д.
11.
Одиночное сокращение1
2
3
1 – латентный период, 2 – фаза сокращения, 3 – фаза
расслабления
Профессор Берг М.Д.
12.
Суммация сокращенийНеполная суммация
Полная суммация
Зубчатый тетанус
Профессор Берг М.Д.
Гладкий тетанус
13.
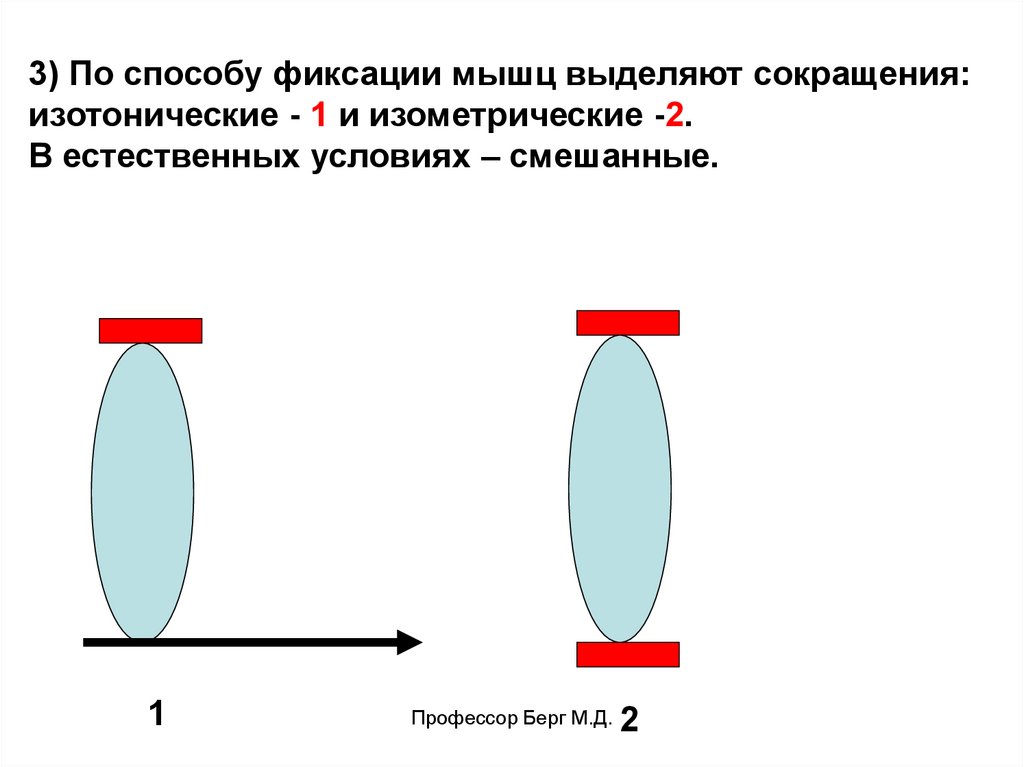
3) По способу фиксации мышц выделяют сокращения:изотонические - 1 и изометрические -2.
В естественных условиях – смешанные.
1
Профессор Берг М.Д.
2
14.
Оценка сократимости1. Относительная сила мышцы, кг (динамометрия)
2. Абсолютная сила мышцы, кг/см2
3. Индекс относительной силы,
кг * 100%:mтела в кг
Профессор Берг М.Д.
15. Механизм сокращения и расслабления мышц
Мышца сокращается, если получает нервныеимпульсы (ПДн).
В эксперименте ее можно раздражать импульсами
электрического тока.
Нервные импульсы с мотонейрона передаются на
мышцу с помощью нервно-мышечного синапса.
В результате мышца возбуждается и сокращается.
Профессор Берг М.Д.
16.
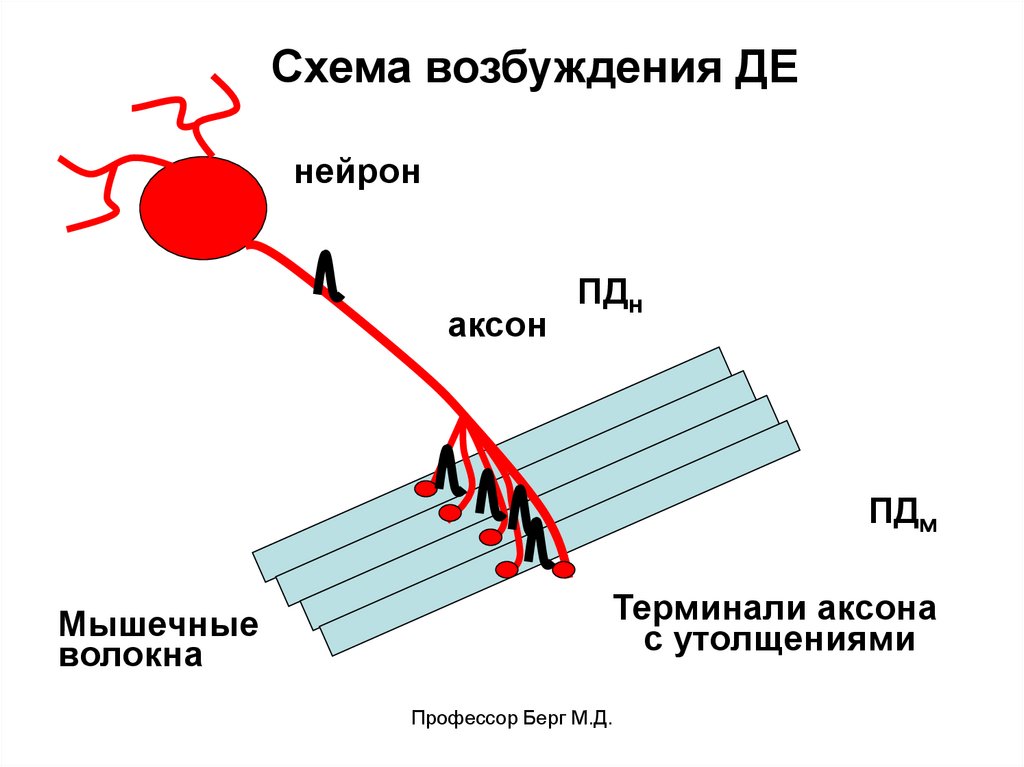
Схема возбуждения ДЕнейрон
аксон
ПДн
ПДм
Мышечные
волокна
Терминали аксона
с утолщениями
Профессор Берг М.Д.
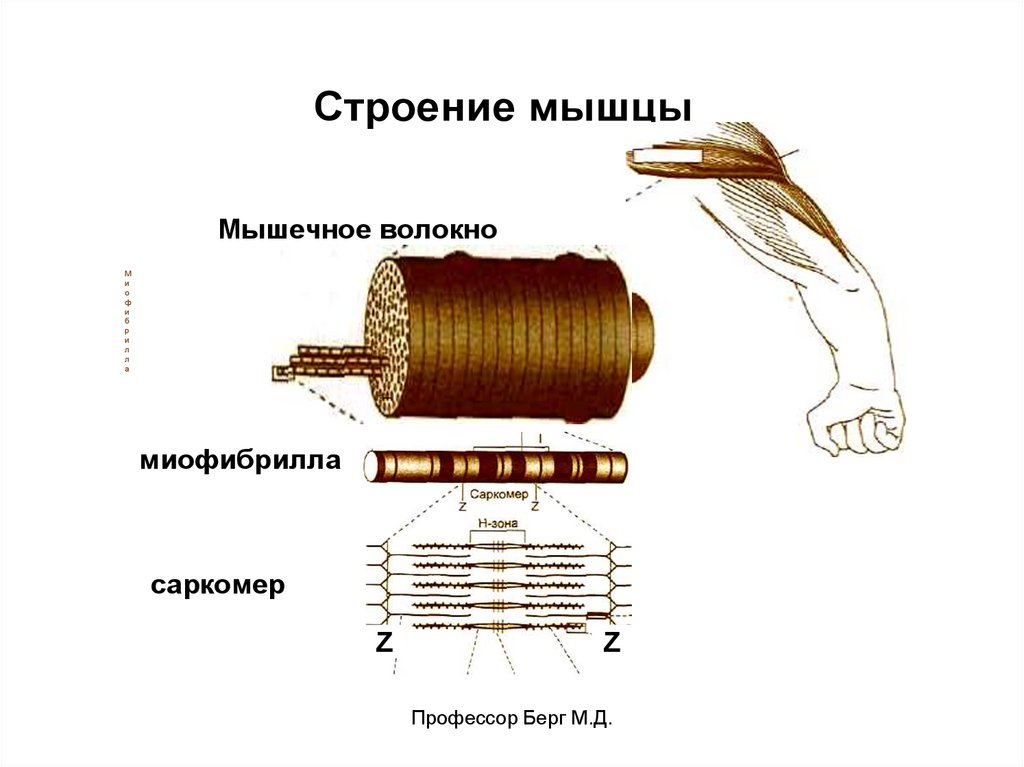
17.
Строение мышцыМышечное волокно
М
и
о
ф
и
б
р
и
л
л
а
-—.
миофибрилла
саркомер
Z
Z
Профессор Берг М.Д.
18.
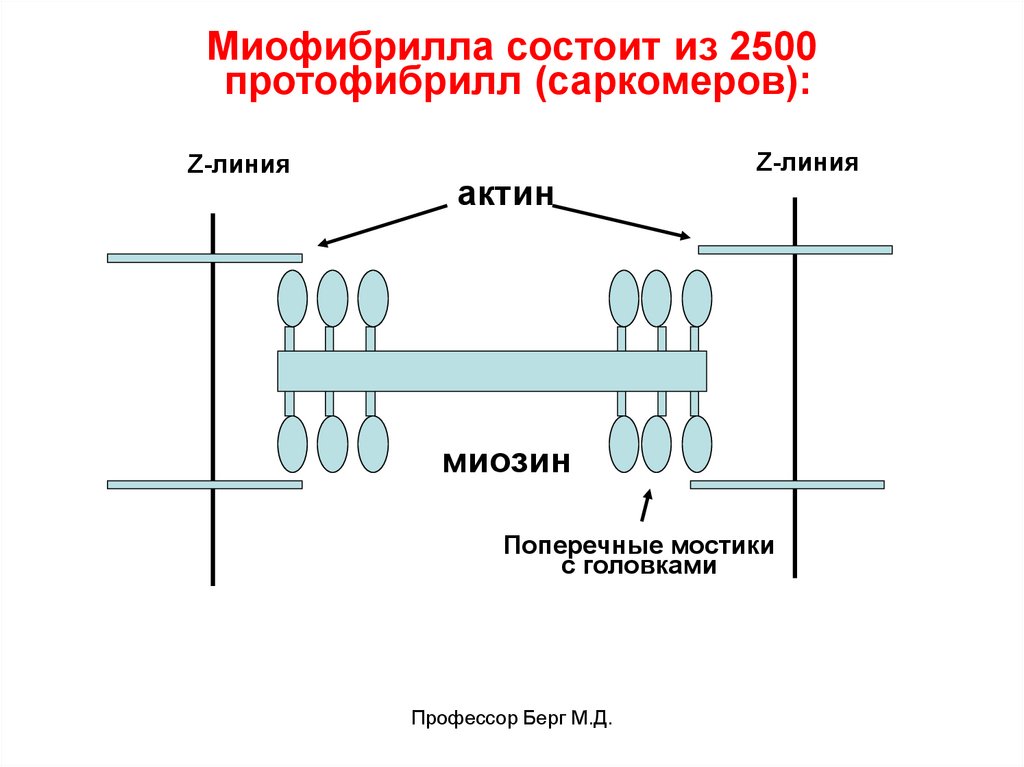
Миофибрилла состоит из 2500протофибрилл (саркомеров):
Z-линия
актин
Z-линия
миозин
Поперечные мостики
с головками
Профессор Берг М.Д.
19.
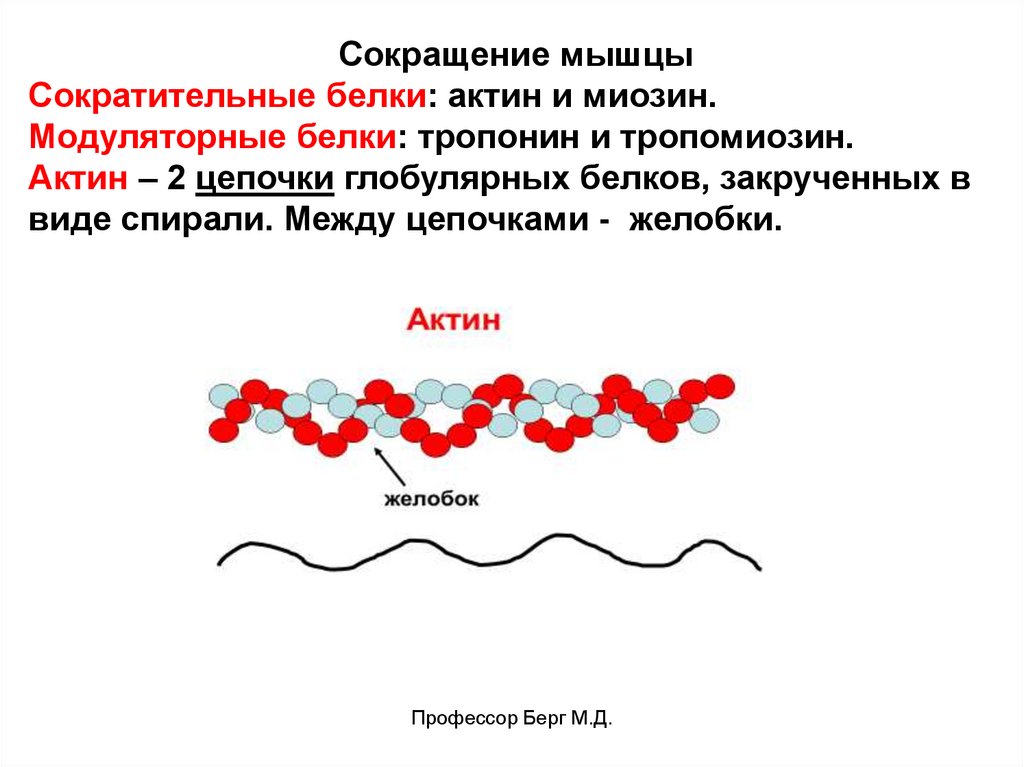
Сокращение мышцыСократительные белки: актин и миозин.
Модуляторные белки: тропонин и тропомиозин.
Актин – 2 цепочки глобулярных белков, закрученных в
виде спирали. Между цепочками - желобки.
Профессор Берг М.Д.
20.
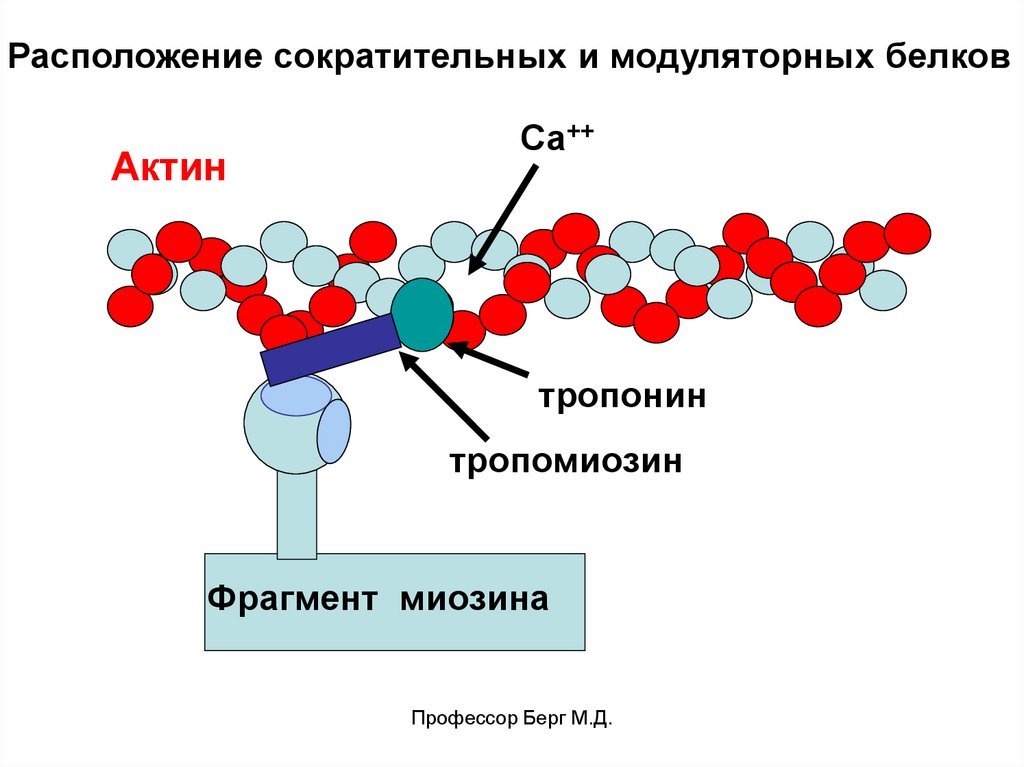
В желобке лежит молекула тропонина, соединенная смолекулой тропомиозина. Тропомиозин закрывает
доступ головок миозина к актину.
Тропонин – глобулярный белок, чувствительный к
ионам Са++. Связываясь с Са++, он изменяет свою
форму и тем самым сдвигает молекулу тропомиозина в
глубину желобка актина. При этом открываются центры
актина для связывания с головкой миозина.
Миозин – нитевидный белок, имеет поперечные
мостики, которые заканчиваются головками миозина.
Головка имеет 2 центра – для связи с актином и для
расщепления АТФ (миозиновая АТФ-аза).
Профессор Берг М.Д.
21.
Расположение сократительных и модуляторных белковАктин
Са++
тропонин
тропомиозин
Фрагмент миозина
Профессор Берг М.Д.
22. Сопряжение возбуждения и сокращения
Профессор Берг М.Д.23.
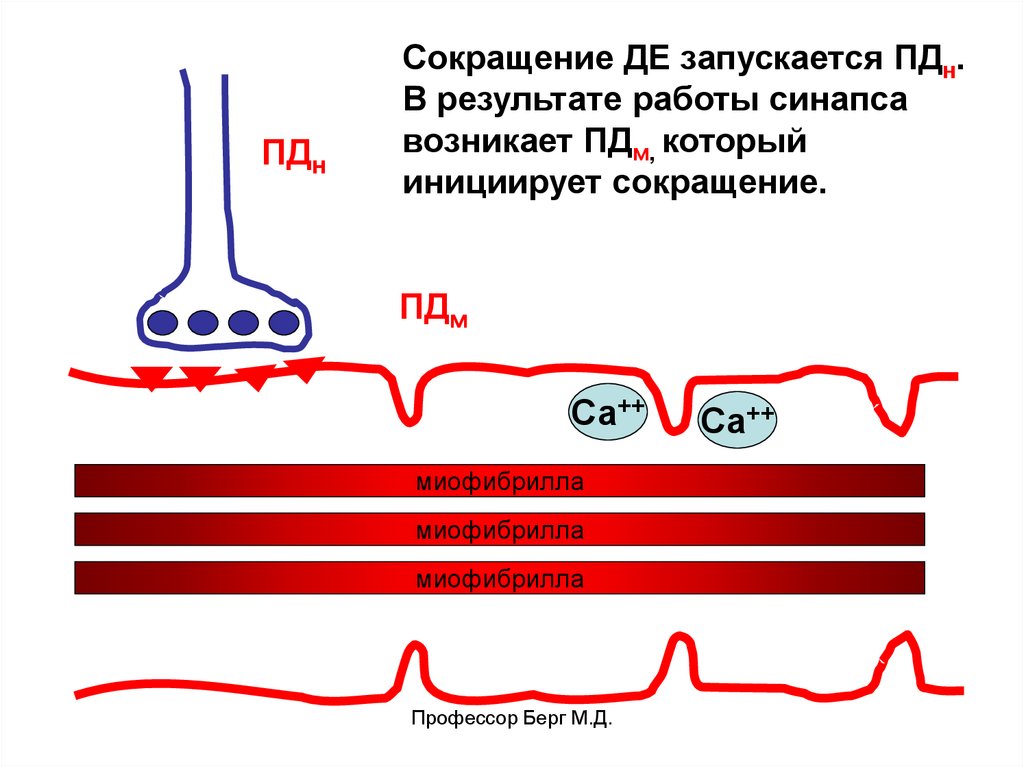
ПДнСокращение ДЕ запускается ПДн.
В результате работы синапса
возникает ПДм, который
инициирует сокращение.
ПДм
Са++
миофибрилла
миофибрилла
миофибрилла
Профессор Берг М.Д.
Са++
24.
Само сокращение запускается ионами Са++.Они являются посредником между возбуждением и
сокращением мышцы.
Ионы Са++ хранятся в цистернах саркоплазматического
ретикулума, расположенных вблизи впячиваний
плазматической мембраны (триады). Са++ хранится в
связанном виде с белком кальсеквестрином. Он
освобождается из цистерн под влиянием ПДм.
Этот процесс называют сопряжением между
возбуждением и сокращением.
Концентрация кальция в цитоплазме повышается при
этом в 100 раз, с 10-7 до 10-5.
Профессор Берг М.Д.
25.
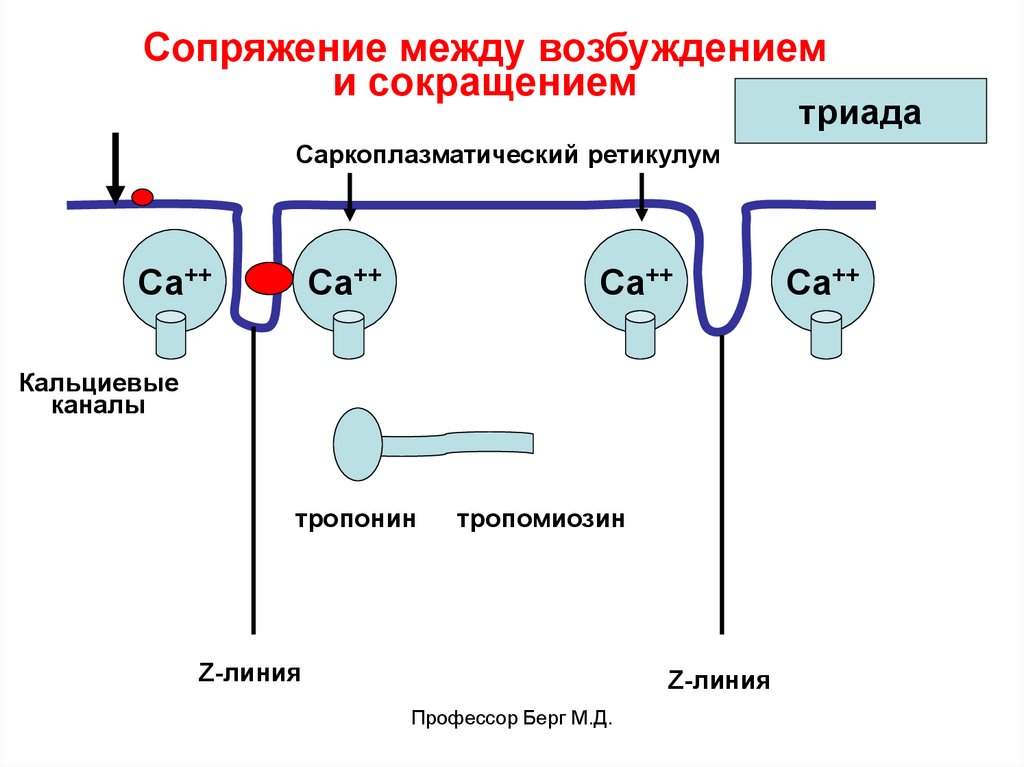
Сопряжение между возбуждениеми сокращением
триада
Саркоплазматический ретикулум
Са++
Са++
Са++
Кальциевые
каналы
тропонин
тропомиозин
Z-линия
Z-линия
Профессор Берг М.Д.
Са++
26.
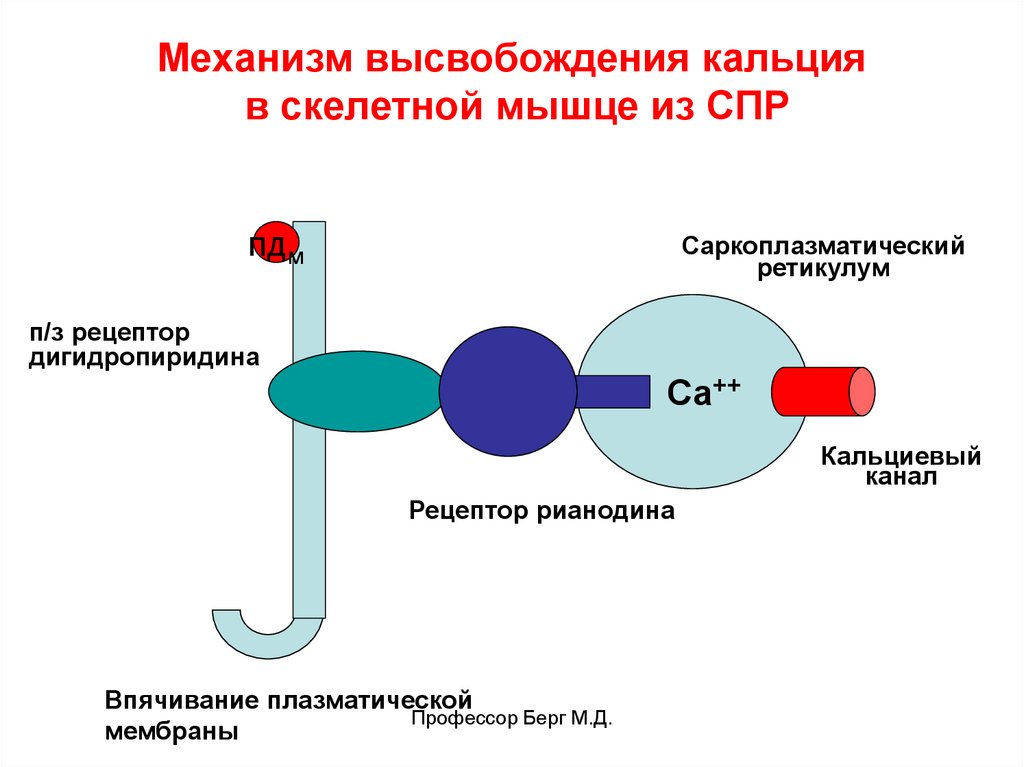
Механизм высвобождения кальцияв скелетной мышце из СПР
пдм
Саркоплазматический
ретикулум
п/з рецептор
дигидропиридина
Са++
Кальциевый
канал
Рецептор рианодина
Впячивание плазматической
Профессор Берг М.Д.
мембраны
27.
Этапность процесса сокращения:1. Распространение ПДм по мембране
мышечного волокна.
2. Выход кальция из депо.
3. Взаимодействие кальция с тропонином.
4. Конформация молекулы тропонина и
смещение молекулы тропомиозина вглубь
желобка актина.
5. Открытие активных центров актина и контакт
с ними головок миозина.
6. Освобождение энергии АТФ (миозиновая
АТФ-аза), сгибание головки миозина и протягивание нити актина вдоль миозина (гребок),
а также спиральное скручивание стромы
мышечного волокна (преодоление эластических
сил).
Профессор Берг М.Д.
28.
7. Присоединение к головке миозина АТФ, отделение ивосстановление исходного положения головки
(нарушение при трупном окоченении).
Этот цикл называют гребковым движением.
Оно протягивает актин вдоль миозина (сокращение),
часть энергии запасается в эластических структурах
для последующего расслабления.
На рабочий цикл расходуется 1 молекула АТФ.
Каждая головка совершает около 5 циклов в сек.
Следующие друг за другом гребковые движения
подтягивают нити актина к центру саркомера, Z-линии
сближаются, мышца укорачивается.
Профессор Берг М.Д.
29.
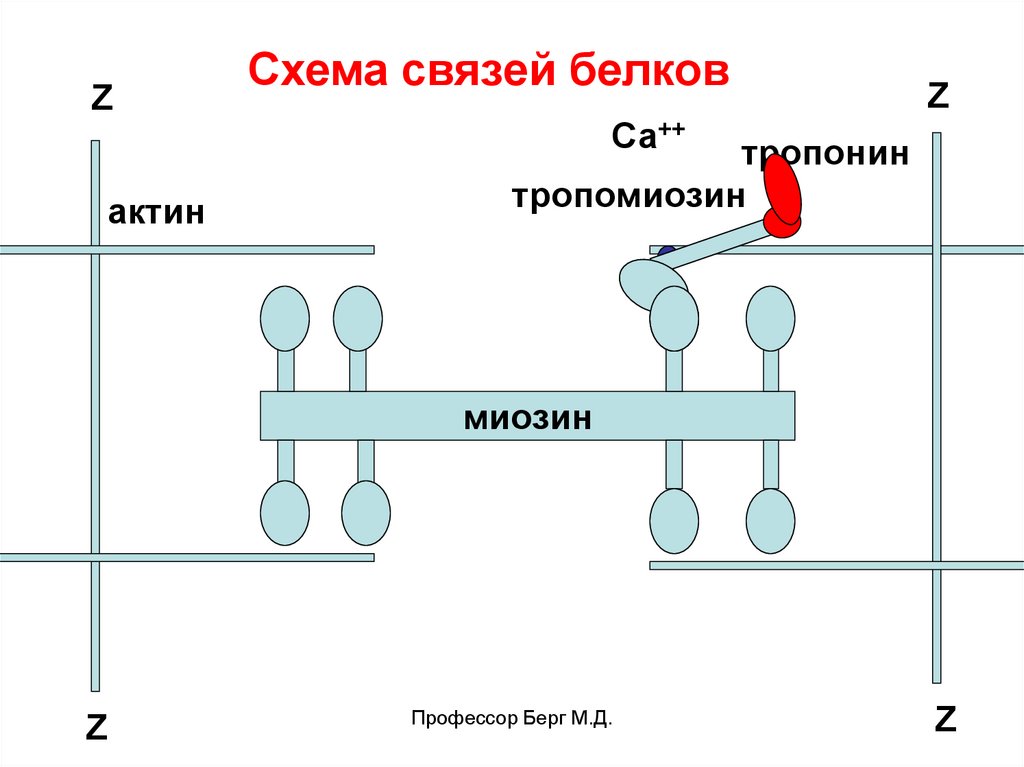
Zактин
Схема связей белков
Z
Са++
тропонин
тропомиозин
миозин
Z
Профессор Берг М.Д.
Z
30.
Молекулы АТФ для сокращения образуются вмитохондриях в процессе окислительного
фосфорилирования в присутствии О2.
Кислород доставляется кровью, а также частично
депонируется в миоглобине (Мb).
Основными источниками энергии в митохондриях
скелетных мышц являются углеводы (глюкоза
и гликоген) и жирные кислоты (ЖК).
В бескислородной среде источником АТФ является
анаэробный гликолиз, а также перенос фосфата от
креатинфосфата на АДФ с образованием АТФ и
креатина. Запаса энергии для работы хватает на:
1. Креатинфосфат (10-12 с).
2. Анаэробный гликолиз (до 1,5-2 мин).
3. Аэробный гликолиз (со 2-й мин и бесконечно).
Профессор Берг М.Д.
31.
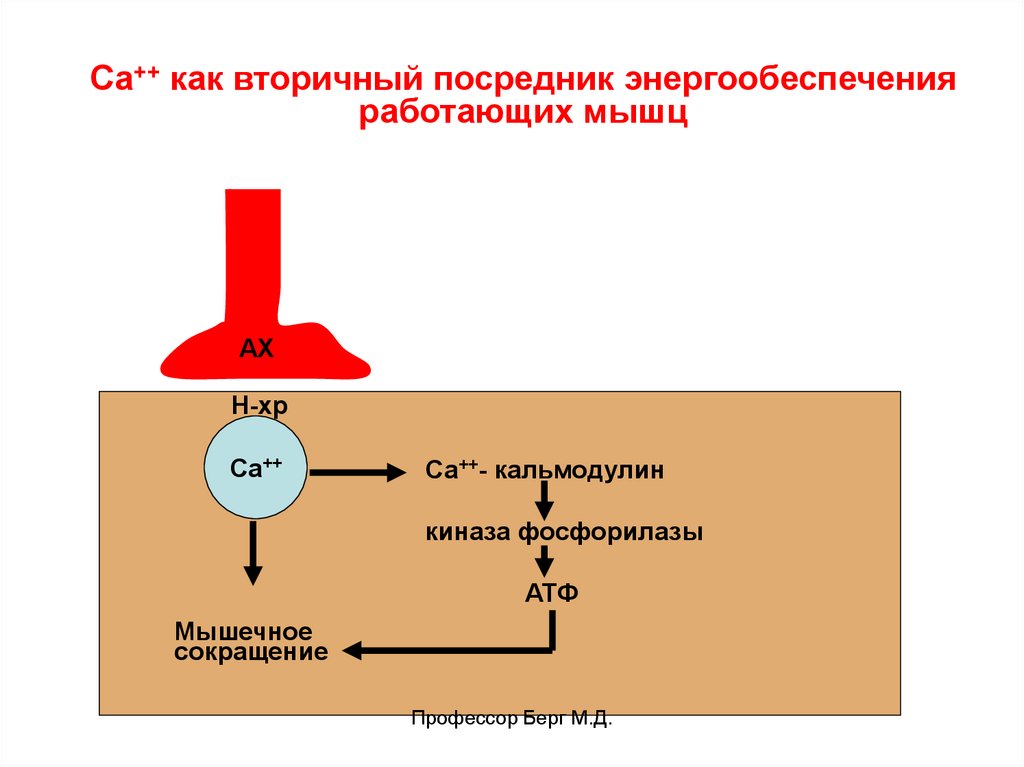
Са++ как вторичный посредник энергообеспеченияработающих мышц
АХ
Н-хр
Са++
Са++- кальмодулин
киназа фосфорилазы
АТФ
Мышечное
сокращение
Профессор Берг М.Д.
32.
Расслабление мышцы.Начинается, если не поступают нервные импульсы.
Повышенная концентрация Са++ в цитоплазме
включает кальциевые насосы СПР, которые
закачивают Са++ против градиента концентрации
обратно в депо – СПР.
Снижение концентрации Са++ в цитоплазме приводит к
отделению его от тропонина, тропомиозин занимает
исходное положение – перекрывает активные центры
актина.
В результате - головки миозина не имеют доступа к
актину.
За счет эластических сил мышца растягивается в
исходное положение – расслабление мышцы.
Профессор Берг М.Д.
33.
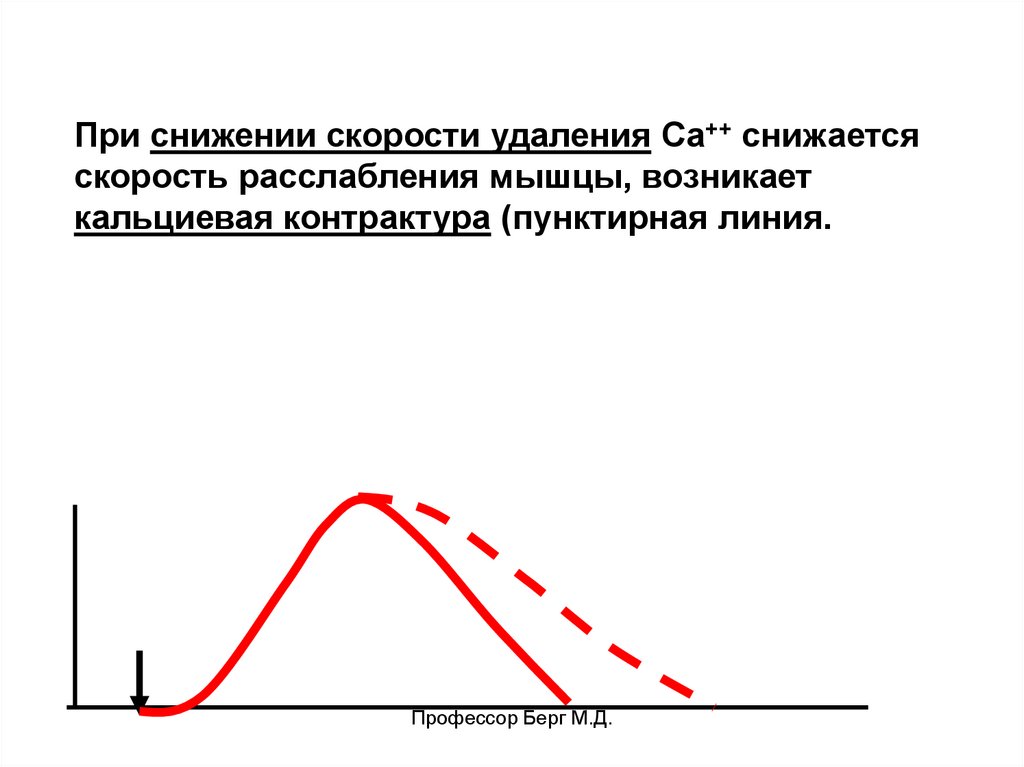
При снижении скорости удаления Са++ снижаетсяскорость расслабления мышцы, возникает
кальциевая контрактура (пунктирная линия.
Профессор Берг М.Д.
34. Физиология гладких мышц
Профессор Берг М.Д.35.
Функции – передвижение жидкостей в организме,формирование объема полых органов, создание
давления в кровеносных и лимфатических сосудах.
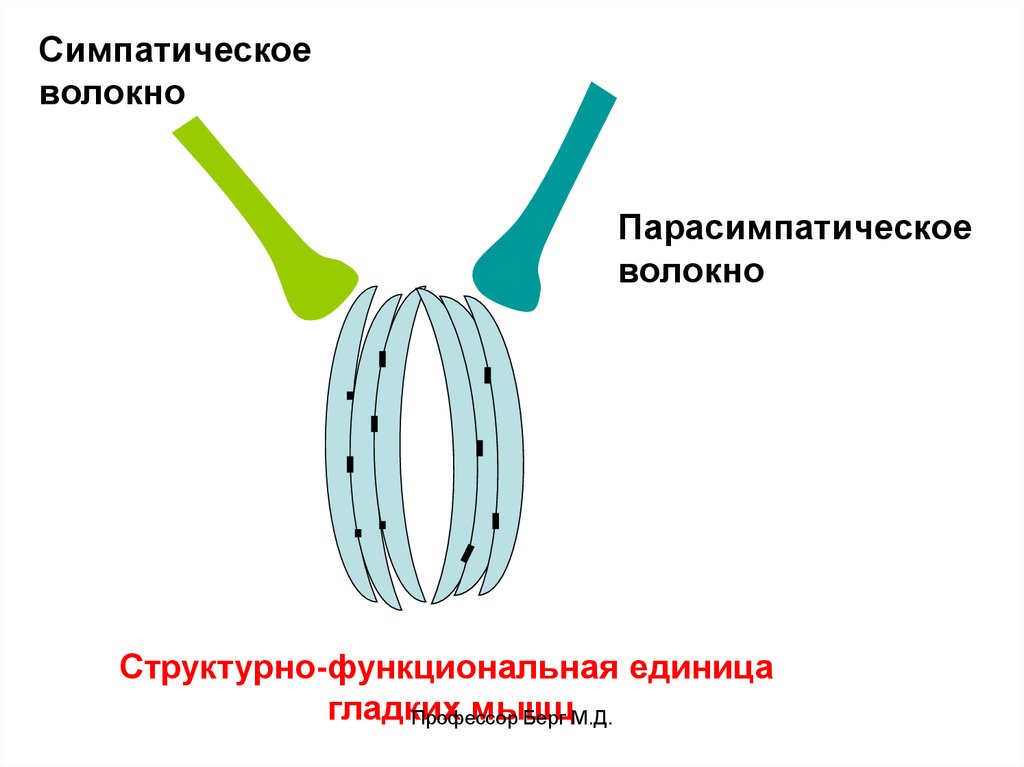
Структурно-функциональная единица – пучок волокон
диаметром 100 мкм.
Естественные раздражители:
• ПД вегетативных нервов,
• гуморальные вещества (гормоны, БАВ),
• быстрое растяжение.
Иннервация: отдельные пучки иннервированы двумя
нервными волокнами – симпатическими и
парасимпатическими, которые образуют вблизи пучка
варикозные расширения, из которых выделяется
медиатор.
Профессор Берг М.Д.
36.
Симпатическоеволокно
Парасимпатическое
волокно
Структурно-функциональная единица
гладких
мышц
Профессор
Берг М.Д.
37.
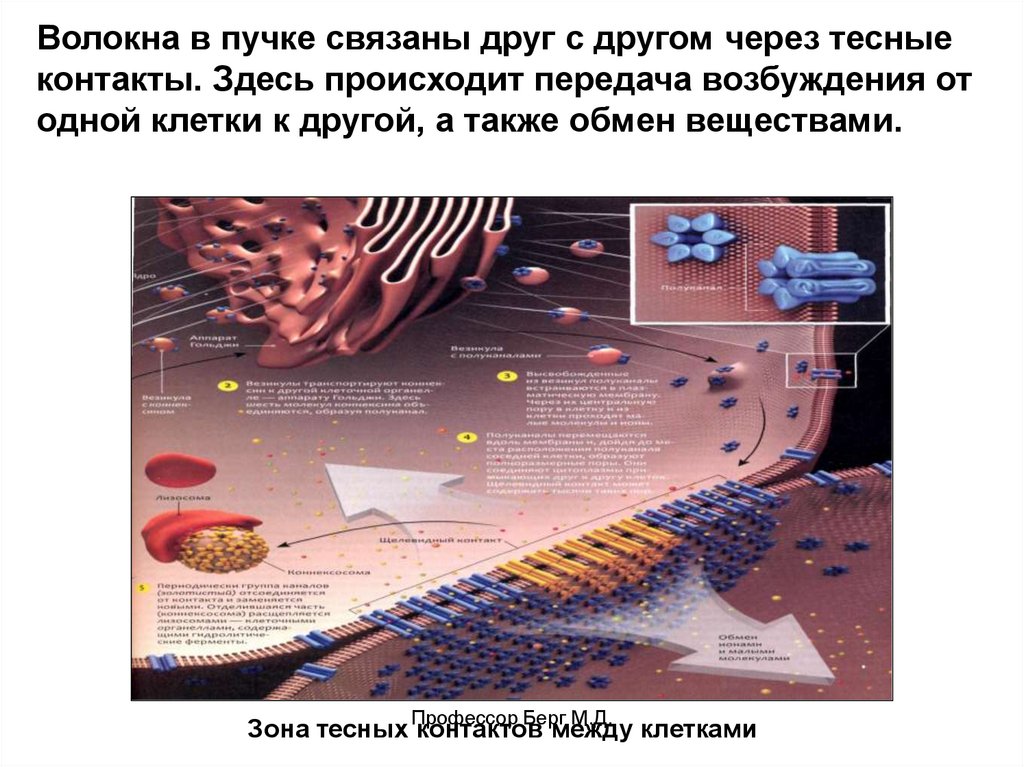
Волокна в пучке связаны друг с другом через тесныеконтакты. Здесь происходит передача возбуждения от
одной клетки к другой, а также обмен веществами.
Берг М.Д.
Зона тесных Профессор
контактов
между клетками
38.
Особенности сокращения:1. Сократительный аппарат не образует типичных
миофибрилл:
• актиновые нити не упорядочены по продольной оси
волокна,
• миозиновые нити расположены между ними,
активация их поперечных мостиков происходит под
влиянием ионов Са++, связанных с кальмодулином.
2. Са++ используется из разных источников:
а) СПР,
б) подмембранные пузырьки,
в) поступает из вне.
Профессор Берг М.Д.
39.
Кальций освобождается под влиянием ПД илихимического стимула, связывается с кальмодулином
(модуляторный белок), этот комплекс активирует
миозинкиназу поперечных мостиков, в результате
головка мостика прикрепляется к актину и долго
удерживается в этом положении при малом
расходовании энергии АТФ.
Механизм гребкового движения такой же, как у п/п
мышцы. На это расходуется 1 молекула АТФ.
Профессор Берг М.Д.
40.
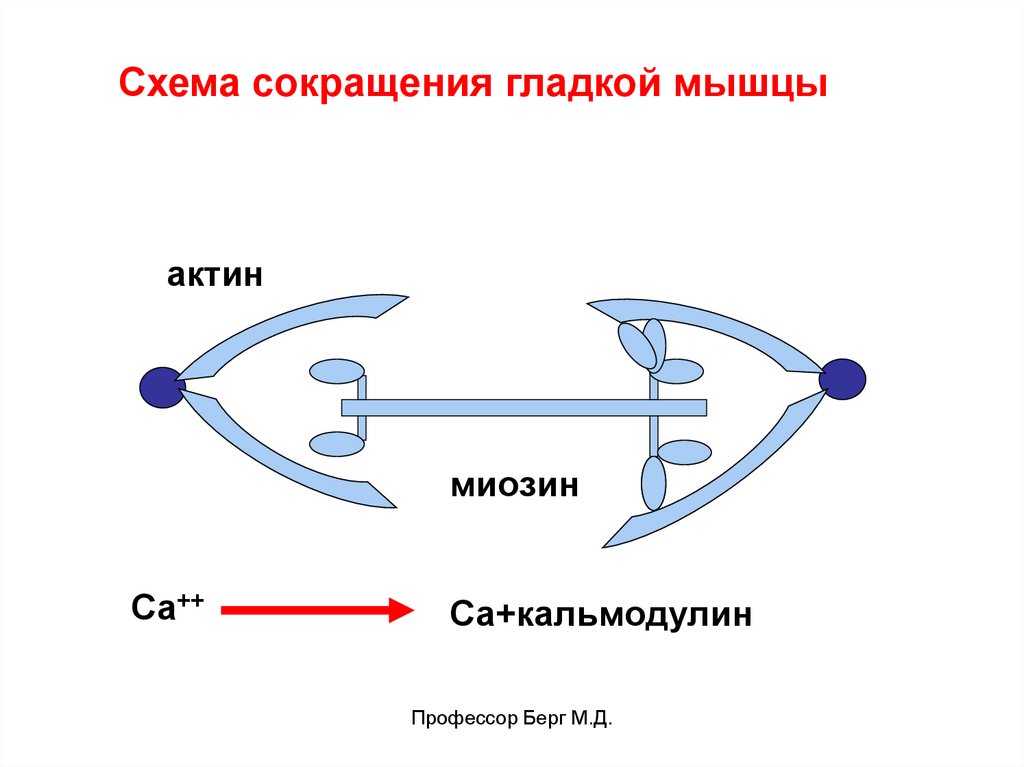
Схема сокращения гладкой мышцыактин
миозин
Са++
Са+кальмодулин
Профессор Берг М.Д.
41.
При снижении концентрации Са++ в цитоплазмепроцесс развивается в обратном направлении
(расслабление).
Для этого кальций закачивается в депо или удаляется
во внеклеточную среду.
Это выполняют кальциевые насосы мембран СПР или
плазматической мембраны.
Профессор Берг М.Д.
42.
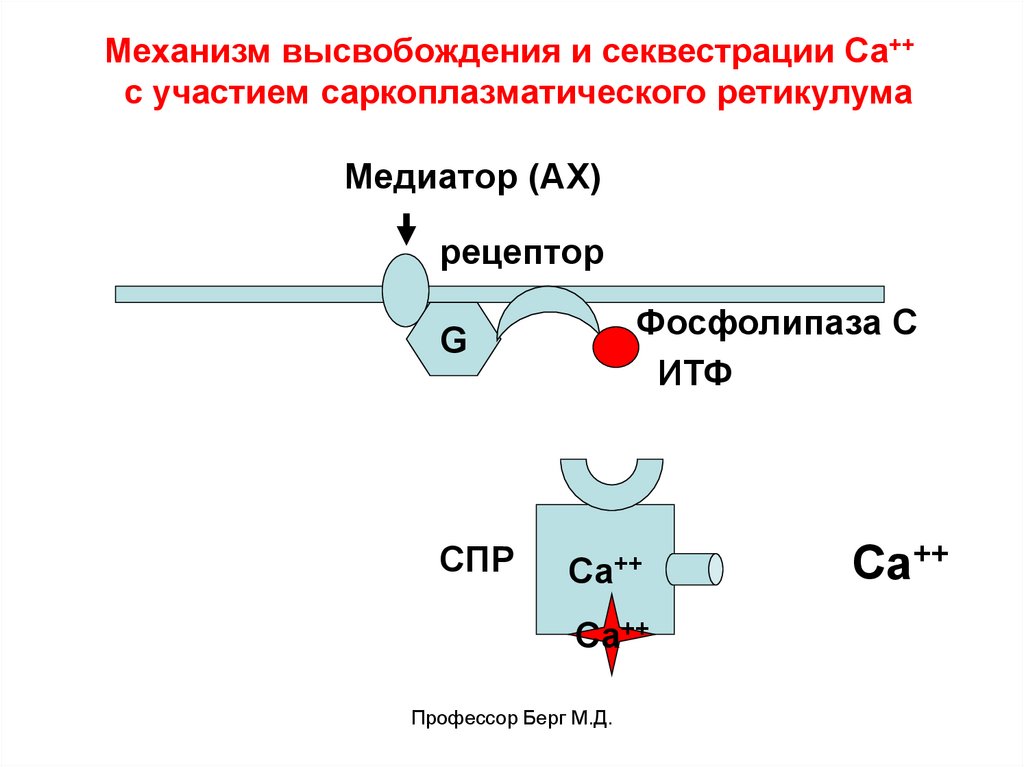
Механизм высвобождения и секвестрации Са++с участием саркоплазматического ретикулума
Медиатор (АХ)
рецептор
Фосфолипаза С
ИТФ
G
СПР
Са++
Са++
Профессор Берг М.Д.
Са++
43.
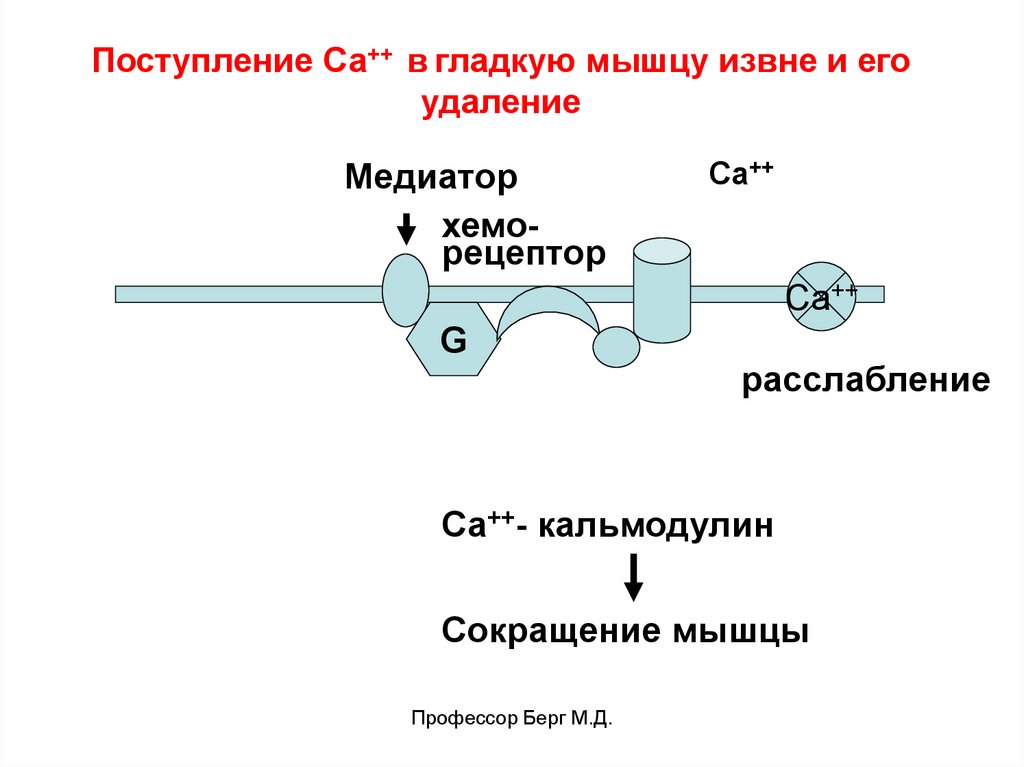
Поступление Са++ в гладкую мышцу извне и егоудаление
Медиатор
хеморецептор
Ca++
Са++
G
расслабление
Са++- кальмодулин
Сокращение мышцы
Профессор Берг М.Д.
44.
Виды сокращений гладких мышц:1.Тонические:
А) базальный тонус. Задается пейсмекерами,
Б) нейрогенный тонус (СС, ПС),
В) гуморальный тонус.
2. Пластический тонус (при медленном растяжении
сохраняется исходный тонус, давление жидкости в
полом органе не меняется (мочевой пузырь).
3. Ритмические сокращения:
а) одиночные,
б) тетанические.
Перистальтические волны, маятникообразные
сокращения, ритмическая сегментация, открытие и
закрытие сфинктеров.
Профессор Берг М.Д.
45.
Время сокращения и расслабления гладкой мышцы на1-2 порядка больше, чем у п/п. Сила сокращения
примерно как у п/п мышцы.
Энергии для поддержания сокращения расходуется
мало, это обеспечивает длительное сокращение без
утомления.
Профессор Берг М.Д.
46.
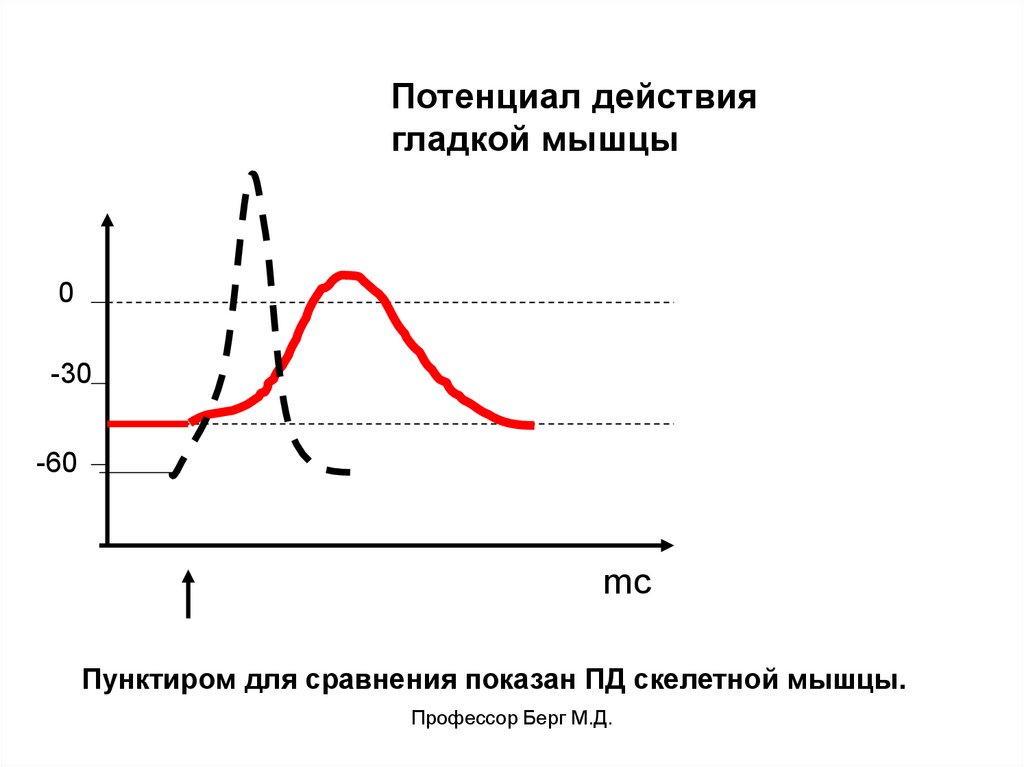
Электрические явленияв гладкой мышце.
ПП = -50-60 мв.
ПД двух видов:
• спайковый. ПД=30-60 мв, длительность – 10-50 мс.
• платообразный. ПД= 50 мв, длительность – 300-1000
мс.
Ионный механизм ПД связан с потенциалзависимыми
Са++ каналами, частично – с Na+ хемозависимыми.
Профессор Берг М.Д.
47.
Потенциал действиягладкой мышцы
0
-30
-60
mс
Пунктиром для сравнения показан ПД скелетной мышцы.
Профессор Берг М.Д.
48.
Автоматия гладких мышцВ определенных участках слоев гладких мышц
располагаются водители ритма (пейсмекеры), т.е.
гладкие мышцы, обладающие автоматией.
Это кардиальный отдел желудка, область сфинктера
Одди ДПК, илеоцекальный сфинктер, прекапиллярные
сфинктеры, лимфангионы лимфатической системы.
Эти клетки обладают спонтанной активностью. Она
связана с метаболизмом клетки, изменением в
цитоплазме концентрации цАМФ и Са++.
Спонтанное увеличение свободного Са++ приводит к
генерации ПД, т.е. к самовозбуждению (автоматия).
Профессор Берг М.Д.
49. Деятельность секреторных клеток
Профессор Берг М.Д.50.
Образуют:• железы внешней секреции
(секрет выделяется в полый орган – соки ЖКТ, пот,
слеза, молоко),
• железы внутренней секреции
(секрет выделяется в кровь – гормоны, местные
гормоны).
Секрет – это плазма крови, обогащенная
секретируемым веществом.
Функционирование секреторной клетки:
• синтез секрета,
• секреция (выделение секрета).
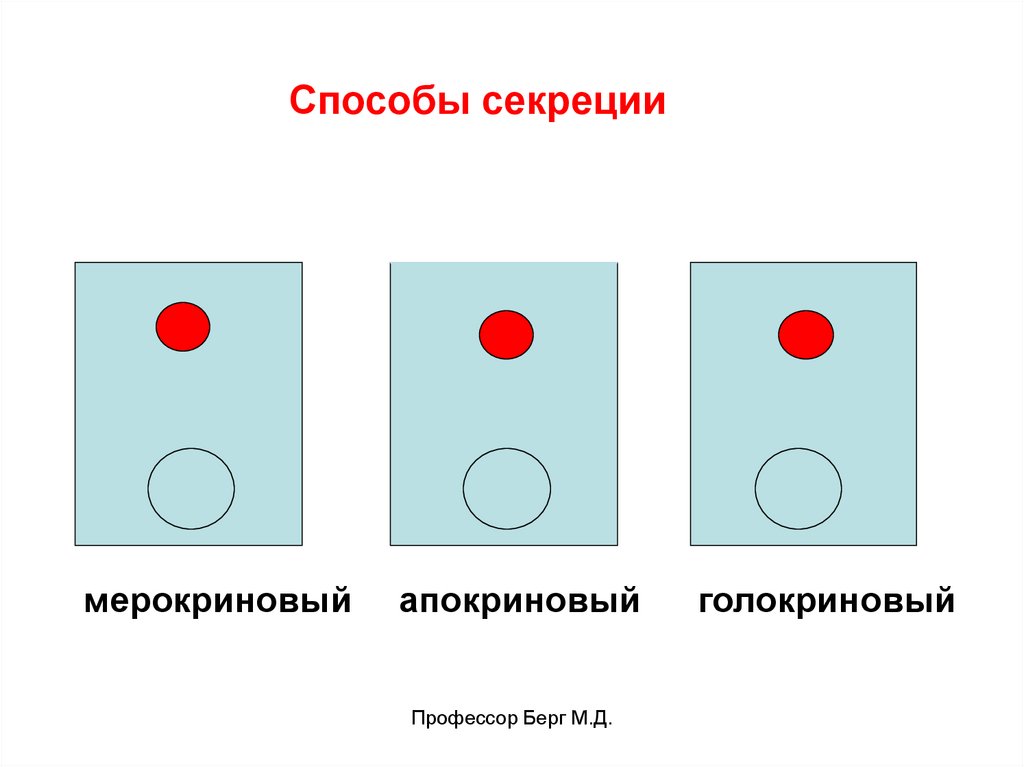
Способы секреции:
1) мерокриновый (экзоцитоз),
2) апокриновый,
3) голокриновый.
Профессор Берг М.Д.
51.
Способы секрециимерокриновый
апокриновый
Профессор Берг М.Д.
голокриновый
52.
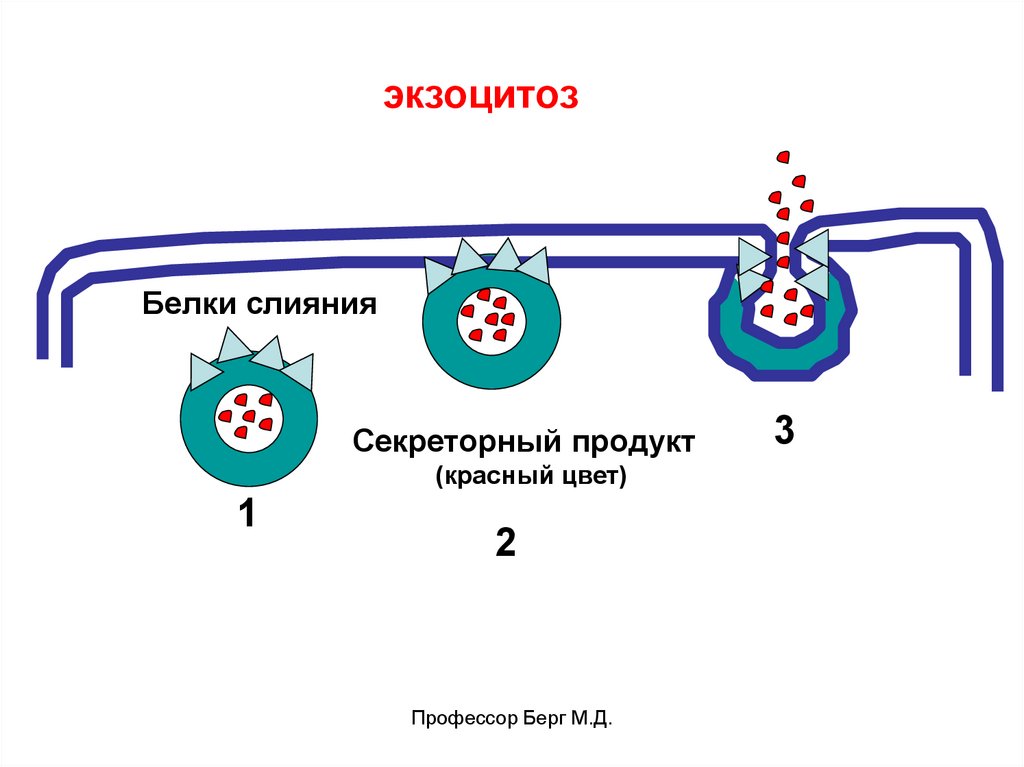
экзоцитозБелки слияния
Секреторный продукт
(красный цвет)
1
2
Профессор Берг М.Д.
3
53.
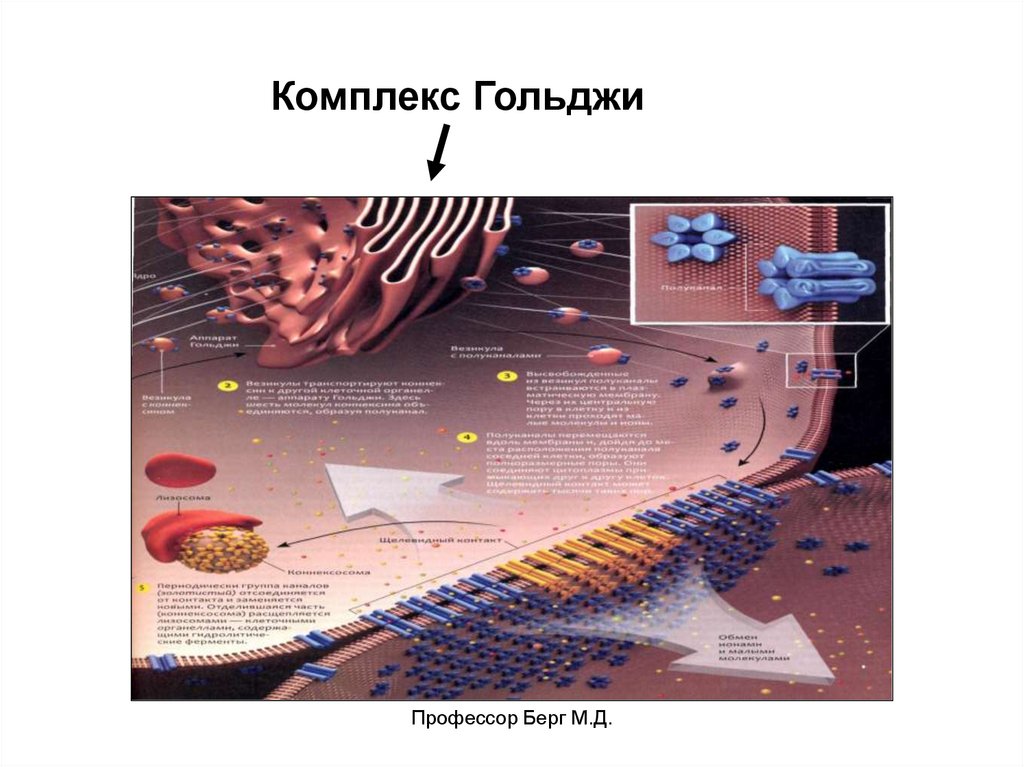
Внутриклеточные структуры синтеза веществ:1. Гранулярная эндоплазматическая сеть
(шороховатый эндоплазматический ретикулум) –
система плоских мембранных цистерн, в которых
происходит синтез белков для экзоцитоза.
2. Комплекс Гольджи – стопка из 3-10 уплощенных
цистерн с расширенными концами.
Здесь происходит модификация секреторного
продукта, сортировка белков, концентрирование и
упаковка секреторных продуктов.
Профессор Берг М.Д.
54.
Комплекс ГольджиПрофессор Берг М.Д.
55.
Виды секреции:1) Базальная (фоновая).
Непрерывная секреция малого объема в состоянии
покоя.
2) Вызванная.
Естественные раздражители вызванной секреции:
• ПД парасимпатической (ПС) или симпатической
системы (СС),
• БАВ,
• Гормоны.
Профессор Берг М.Д.
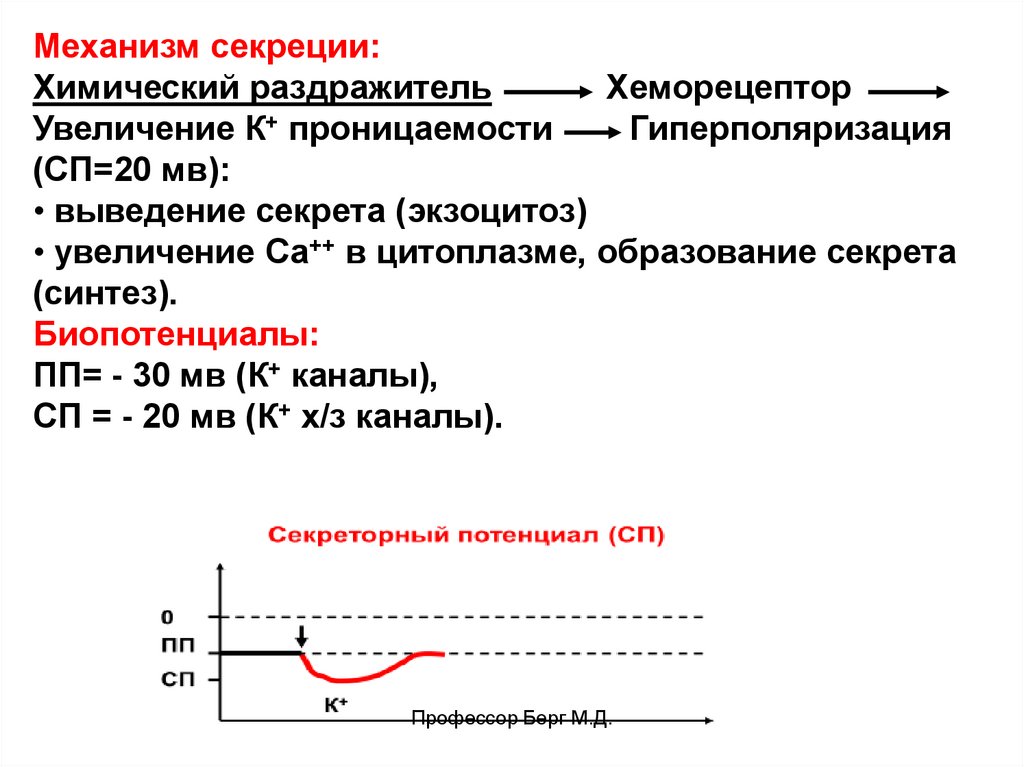
56.
Механизм секреции:Химический раздражитель
Хеморецептор
Увеличение К+ проницаемости
Гиперполяризация
(СП=20 мв):
• выведение секрета (экзоцитоз)
• увеличение Са++ в цитоплазме, образование секрета
(синтез).
Биопотенциалы:
ПП= - 30 мв (К+ каналы),
СП = - 20 мв (К+ х/з каналы).
Профессор Берг М.Д.