


")



 МЫШЕЧНАЯ ТКАНЬ")










 МЫШЕЧНАЯ ТКАНЬ")










 biology
biologySimilar presentations:










Мышечные ткани. Классификация
1.
МЫШЕЧНЫЕТКАНИ
ГРУППА ТКАНЕЙ ХАРАКТЕРИЗУЮЩИХСЯ
ОБЩЕЙ И ОСНОВНОЙ СПОСОБНОСТЬЮ К
СОКРАЩЕНИЮ,
БЛАГОДАРЯ КОТОРОЙ
ПЕРЕМЕЩАЮТ
ТЕЛО ИЛИ ЧАСТИ ЕГО В ПРОСТРАНСТВ
2.
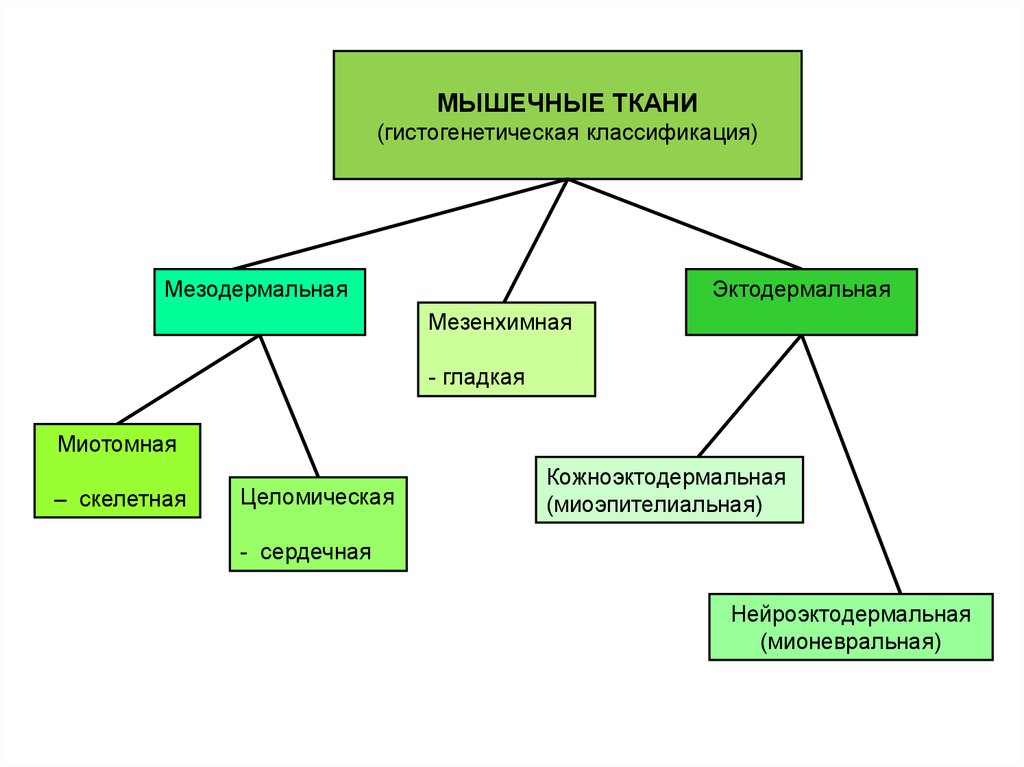
МЫШЕЧНЫЕ ТКАНИ(гистогенетическая классификация)
Мезодермальная
Эктодермальная
Мезенхимная
- гладкая
Миотомная
– скелетная
Целомическая
Кожноэктодермальная
(миоэпителиальная)
- сердечная
Нейроэктодермальная
(мионевральная)
3.
РАЗВИТИЕ МЫШЕЧНОЙ ТКАНИМИОТОМЫСКЕЛЕТНАЯ
МЕЗЕНХИМНАЯГЛАДКАЯ
НЕЙРОЭКТОДЕРМАМИОНЕЙРАЛЬНАЯ
ВИСЦЕРАЛЬНЫЙ
СПЛАНХНОТОМ –
МИОКАРД
4.
Классификация мышечных тканей(по Р.К. Данилову)
МЫШЕЧНЫЕ ТКАНИ
Мезодермальные
1. Спланхномезодермальная
(висцеральная, сосудистая)
2. Миотомная (соматическая,
переднего отдела пищеварительного канала)
3. Целомическая
(кардиомиоциты)
Эктодермальные
1.
Нейральная
(мионейральная)
2. Эктомезенхимная
(дермальная)
5. Классификация миоидных тканей (по Р.К. Данилову)
Миоидные клеткиЭктодермальные
Мезодермальные
Спланхномезенхимные
1. Эктомезенхимные
(миоидные клетки ЦНС)
(миофибробласты,
2. Эпидермальные
миоидные клетки семенных
канальцев,
(миоэпителиальные,
миоидные клетки ЮГ-комплекса,
ренинсекретирующие клетки)
миоидные клетки эпидермиса
3. Прехордальные
(миоидные клетки тимуса
и аденогипофиза)
6.
МЫШЕЧНАЯ ТКАНЬПОПЕРЕЧНОПОЛОСАТАЯ
СКЕЛЕТНАЯ
ПРОИЗВОЛЬНАЯ
ГЛАДКАЯ
СЕРДЕЧНАЯ
НЕПРОИЗВОЛЬНАЯ
7.
У высших позвоночных по генетическим признакам И.Н.Борисов выделяет двегруппы мышечных тканей: целомические (мезодермальные) и нейродермальные.
По морфологическим признакам на 4 вида: поперечнополосатую
миосимпластическую, сердечную, гладкую и пигментную (радужная часть сетчатки, где
преобладают миоэпителиальные клетки, содержащие пигмент).
А.А.Заварзин (1985) предложил делить мышечные ткани многоклеточных на 3
типа. Эта классификация основана на структурной организации сократимого аппарата.
1) поперечно-полосатую и косоисчерченную (с упорядоченным расположением
миофибрилл);
2 )гладкую мускулатуру беспозвоночных;
3) гладкую мускулатуру позвоночных.
По физиологическим особенностям мышечные ткани делят на произвольные и
непроизвольные, а также на тонические (гладкая и сердечная мускулатура) и
тетанические (поперечно-полосатые мышцы).
Различают висцеральную мускулатуру (мышцы внутренних органов) и соматическую
(двигательную).
Наиболее подробная гистогенетическая классификация мышечных тканей предложена
Р.К.Даниловым (1994). Автор, помимо мышечной, выделяет группу клеток, имеющих
большое сходство с мышечными, которые называют миоидными.
Чаще, однако, пользуются классификацией мышечных тканей, выделяющие 3
разновидности:
гладкая, скелетная и сердечная.
8. КРАТКАЯ ХАРАКТЕРИСТИКА ТРЕХ РАЗНОВИДНОСТЕЙ МЫШЕЧНЫХ ТКАНЕЙ
РАСПОЛОЖЕНИЕВ ОРГАНИЗМЕ
МИКРОСТРУКТУРА
НАЛИЧИЕ
МИОФИБРИЛЛ
ПРОИСХОЖДЕНИЕ
ХАРАКТЕР
СОКРАЩЕНИЯ
КЛЕТОЧНАЯ
ГЛАДКАЯ
ВО
ВНУТРЕННИХ
ОРГАНАХ
НЕТ МИОФИБРИЛЛ, ЕСТЬ
ПРОТОФИБРИЛЛЫ
МЕЗЕНХИМА
(СПЛАНХНОМЕЗЕНХИМА)
НЕПРОИЗВОЛЬНЫЙ,
ТОНИЧЕСКИЙ
СКЕЛЕТНАЯ
В
СКЕЛЕТНЫХ
МЫШЦАХ
СИМПЛАСТИЧЕСКАЯ
ЕСТЬ
МИОФИБРИЛЛЫ
ИЗ
МИОТОМОВ
ПРОИЗВОЛЬНЫЙ,
ТЕТАНИЧЕСКИЙ
(РЕДКО
ТОНИЧЕСКИЙ)
В
МИОКАРДЕ
КЛЕТОЧНАЯ
МИОФИБРИЛЛЫ НЕПРАВИЛЬНОЙ
ФОРМЫ
ИЗ
ЦЕЛОМИЧЕСКОЙ
ВЫСТИЛКИ
НЕПРОИЗВОЛЬНЫЙ, ТОНИЧЕСКИЙ, РИТМИЧЕСКИЙ
СЕРДЕЧНАЯ
9. СОМАТИЧЕКАЯ (СКЕЛЕТНАЯ) МЫШЕЧНАЯ ТКАНЬ
ЛОКОМОТОРНЫЕНЕЛОКОМОТОРНЫЕ
(ВИСЦЕРАЛЬНЫЕ)
МЫШЦЫ,
ПРИКРЕПЛЯЮЩИЕСЯ
К СКЕЛЕТУ:
МЫШЕЧНЫЕ
ВОЛОКНА,
СВЯЗАННЫЕ С
МЯГКИМИ ТКАНЯМИ
КОСТЯМ,
СУХОЖИЛИЯМ.
(ВЕРХНИЕ ОТДЕЛЫ
ПИШЕВОДА, ЯЗЫК)
Симпласт. На что бы он похож?
А он какой бы ни был разный,
Напоминает нам колхоз
С обобществленной цитоплазмой.
А.Г. Кнорре
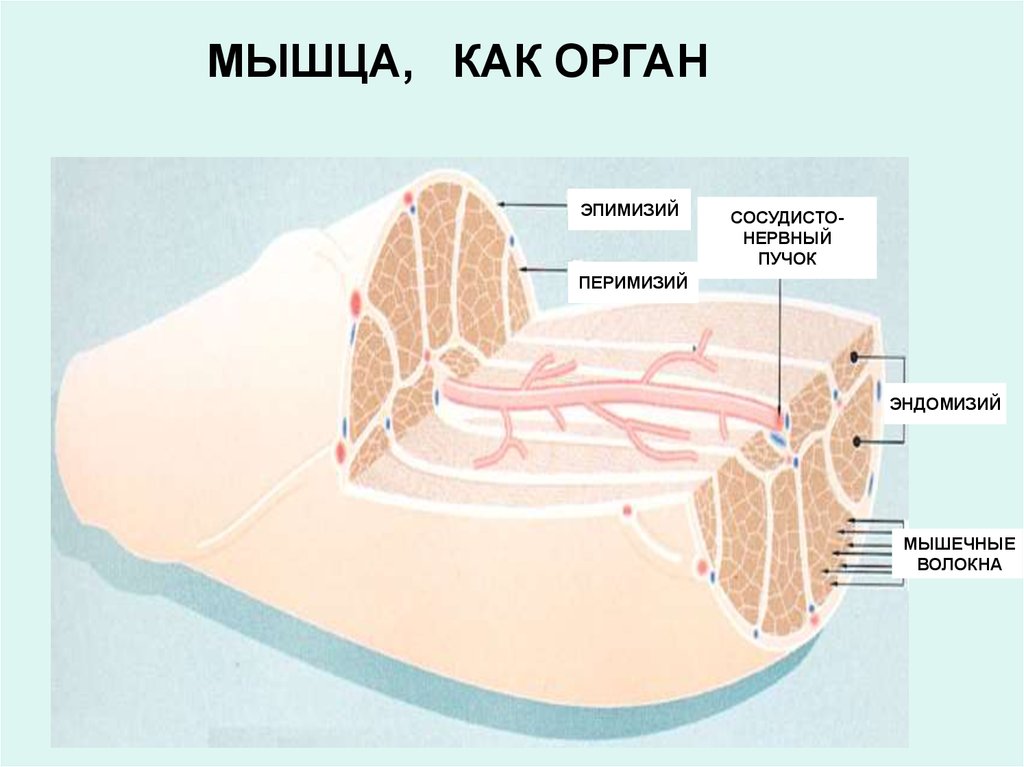
10.
МЫШЦА, КАК ОРГАНЭПИМИЗИЙ
СОСУДИСТОНЕРВНЫЙ
ПУЧОК
ПЕРИМИЗИЙ
ЭНДОМИЗИЙ
МЫШЕЧНЫЕ
ВОЛОКНА
11.
Индивидуальное мышечное волокно окружено плазмолеммой, называемойсарколеммой, которую покрывает базальная мембрана (гликокаликс) толщиной 30-50 нм.
Гликокаликс состоит их фибриллярных структур и аморфного вещества, богатого ГАГ и
протеогликанами. В аморфное вещество впаяны зрелые коллагеновые волокна и
аргирофильные филаменты эндомизия.
Р.К.Данилов под мышечным волокном предлагает понимать образование,
состоящее из двух частей: симпластической и клеточной (миосателлиты), объединенных
общей базальной мембраной, т.е. они представляют собой клеточносимпластические системы, формирующиеся в эмбриональном миогенезе и
сохраняющие свою организацию в постнатальном онтогенезе и после травмы.
12.
ФУНКЦИОНАЛЬНЫЕ АППАРАТЫ МИОСИМПЛАСТА1. СОКРАТИТЕЛЬНЫЙ
(МИОФИБРИЛЛЫ);
2. ПЕРЕДАЧИ ВОЗБУЖДЕНИЯ
(САРКОТУБУЛЯРНАЯ СИСТЕМА);
3. ОПОРНЫЙ
(ЭЛЕМЕНТЫ ЦИТОСКЕЛЕТА, САРКОЛЕММА, БАЗАЛЬНАЯ МЕМБРАНА);
4. ЭНЕРГЕТИЧЕСКИЙ
(МИТОХОНДРИИ, ТРОФИЧЕСКИЕ ВКЛЮЧЕНИЯ);
5. СИНТЕТИЧЕСКИЙ
(РИБОСОМЫ, ПОЛИРИБОСОМЫ,
6. ЛИЗОСОМАЛЬНЫЙ
ГРЭПС,
КОМПЛЕКС ГОЛЬДЖИ);
13. Строение поперечнополосатой мускулатуры
Гликокаликс(базальная мембрана)
САРКОПЛАЗМАТИЧЕСКАЯ СЕТЬ И Т-СИСТЕМЫ
14.
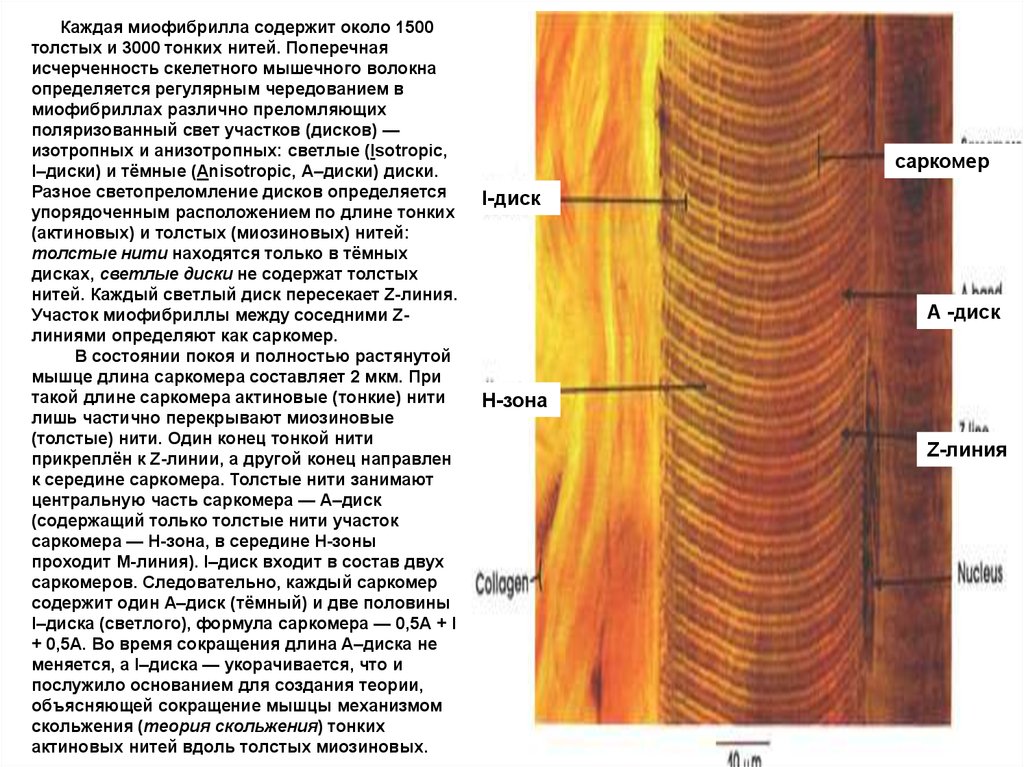
Каждая миофибрилла содержит около 1500толстых и 3000 тонких нитей. Поперечная
исчерченность скелетного мышечного волокна
определяется регулярным чередованием в
миофибриллах различно преломляющих
поляризованный свет участков (дисков) —
изотропных и анизотропных: светлые (Isotropic,
I–диски) и тёмные (Anisotropic, А–диски) диски.
Разное светопреломление дисков определяется
упорядоченным расположением по длине тонких
(актиновых) и толстых (миозиновых) нитей:
толстые нити находятся только в тёмных
дисках, светлые диски не содержат толстых
нитей. Каждый светлый диск пересекает Z-линия.
Участок миофибриллы между соседними Zлиниями определяют как саркомер.
В состоянии покоя и полностью растянутой
мышце длина саркомера составляет 2 мкм. При
такой длине саркомера актиновые (тонкие) нити
лишь частично перекрывают миозиновые
(толстые) нити. Один конец тонкой нити
прикреплён к Z-линии, а другой конец направлен
к середине сaркомера. Толстые нити занимают
центральную часть сaркомера — А–диск
(содержащий только толстые нити участок
сaркомера — Н-зона, в середине Н-зоны
проходит М-линия). I–диск входит в состав двух
сaркомеров. Следовательно, каждый сaркомер
содержит один А–диск (тёмный) и две половины
I–диска (светлого), формула саркомера — 0,5А + I
+ 0,5А. Во время сокращения длина A–диска не
меняется, а I–диска — укорачивается, что и
послужило основанием для создания теории,
объясняющей сокращение мышцы механизмом
скольжения (теория скольжения) тонких
актиновых нитей вдоль толстых миозиновых.
саркомер
I-диск
А -диск
H-зона
Z-линия
15. Строение молекулы миозина
СТЕРЖЕНЬТОЛСТОЙ НИТИ
ХВОСТОВЫЕ НИТИ МИОЗИНА
ГОЛОВКИ МИОЗИНА
S-субфрагмент
ГОЛОВКИ
ЛЕГКИЕ ЦЕПИ
АГРЕГАЦИЯ
ШАРНИРНЫЙ
ШАРНИРНЫЙ
УЧАСТОК
УЧАСТОК
МОЛЕКУЛЫ
ЭЛАСТИЧЕСКИЙ КОМПАНЕНТ
МИОЗИНА
ТЯЖЕЛЫЙ МЕРОМИОЗИН
SГлобулярная
1
головка миозина (S1)
МОЛЕКУЛЫ
Sэластический
2
компонент
субфрагмента
(S
2)
ШАРНИРНЫЙ УЧАСТОК
МИОЗИНА
ЛЕГКИЙ МЕРОМИОЗИН
СВЯЗЫВАЕТ КОНЦЫ ТОЛСТЫХ НИТЕЙ С Z
ЛИНИЕЙ БЕЛОК- ТИТИН;
ОБЪЕДИНЯЕТ ТОЛСТЫЕ И ТОНКИЕ НИТИ
БЕЛОК – НЕБУЛИН.
МИОМЕЗИН (М-БЕЛОК) - СВЯЗЫВАЕТ И
ОРГАНИЗУЕТ ТОЛСТЫЕ НИТИ.
16.
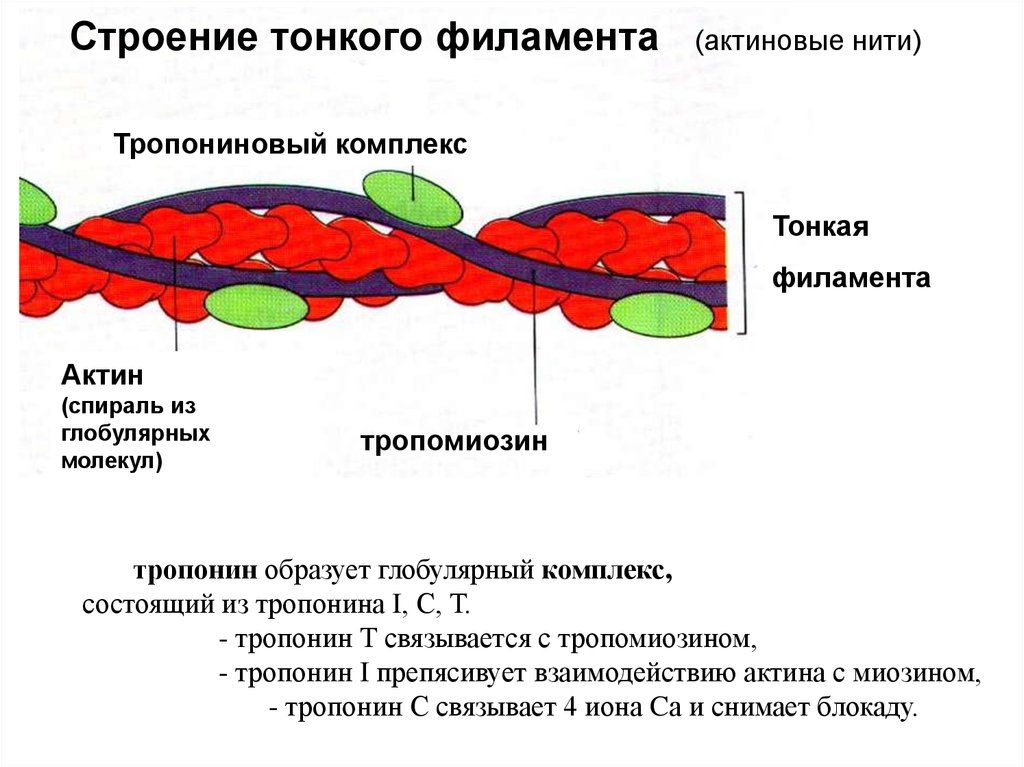
Строение тонкого филамента(актиновые нити)
Тропониновый комплекс
Тонкая
филамента
Актин
(спираль из
глобулярных
молекул)
тропомиозин
тропонин образует глобулярный комплекс,
состоящий из тропонина I, С, Т.
- тропонин Т связывается с тропомиозином,
- тропонин I препясивует взаимодействию актина с миозином,
- тропонин С связывает 4 иона Cа и снимает блокаду.
17.
А-дискБелки М-линии
М-линия
ядро
I-диск
Z-линия
М-линия
изозим креатиназа
(является преобразователем
АДФ в АТФ),
М-белок, миомезин являются местом
прикрепления цитоскелета.
18.
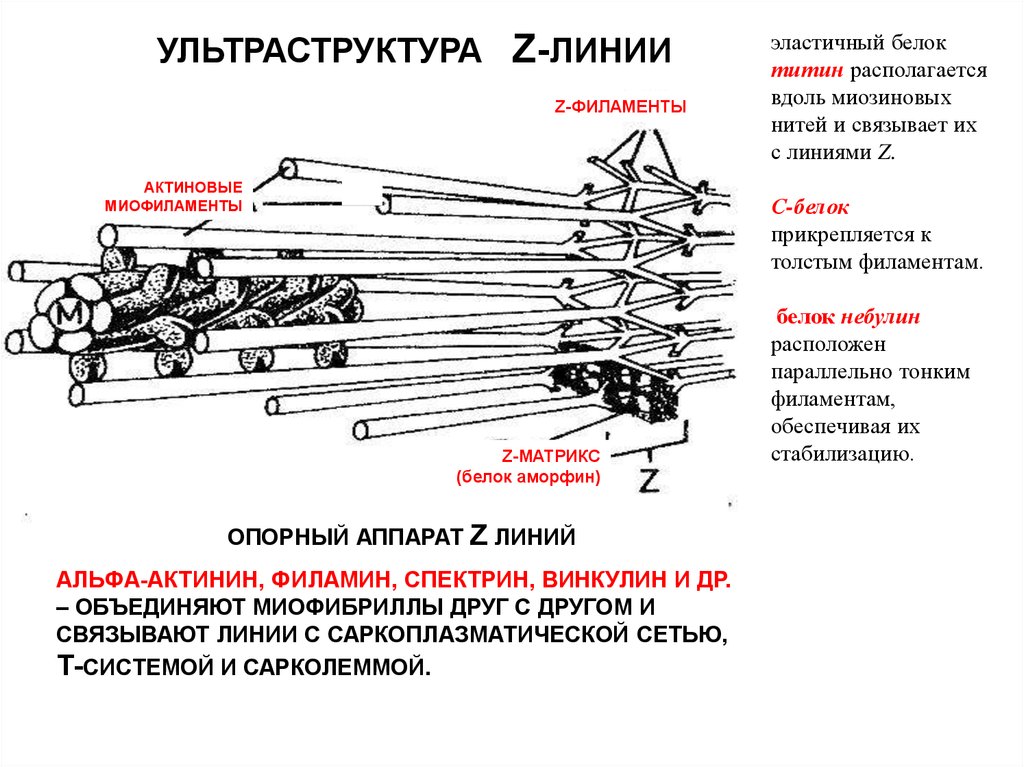
УЛЬТРАСТРУКТУРА Z-ЛИНИИZ-ФИЛАМЕНТЫ
АКТИНОВЫЕ
МИОФИЛАМЕНТЫ
эластичный белок
титин располагается
вдоль миозиновых
нитей и связывает их
с линиями Z.
С-белок
прикрепляется к
толстым филаментам.
Z-МАТРИКС
(белок аморфин)
ОПОРНЫЙ АППАРАТ
Z ЛИНИЙ
АЛЬФА-АКТИНИН, ФИЛАМИН, СПЕКТРИН, ВИНКУЛИН И ДР.
– ОБЪЕДИНЯЮТ МИОФИБРИЛЛЫ ДРУГ С ДРУГОМ И
СВЯЗЫВАЮТ ЛИНИИ С САРКОПЛАЗМАТИЧЕСКОЙ СЕТЬЮ,
Т-СИСТЕМОЙ И САРКОЛЕММОЙ.
белок небулин
расположен
параллельно тонким
филаментам,
обеспечивая их
стабилизацию.
19.
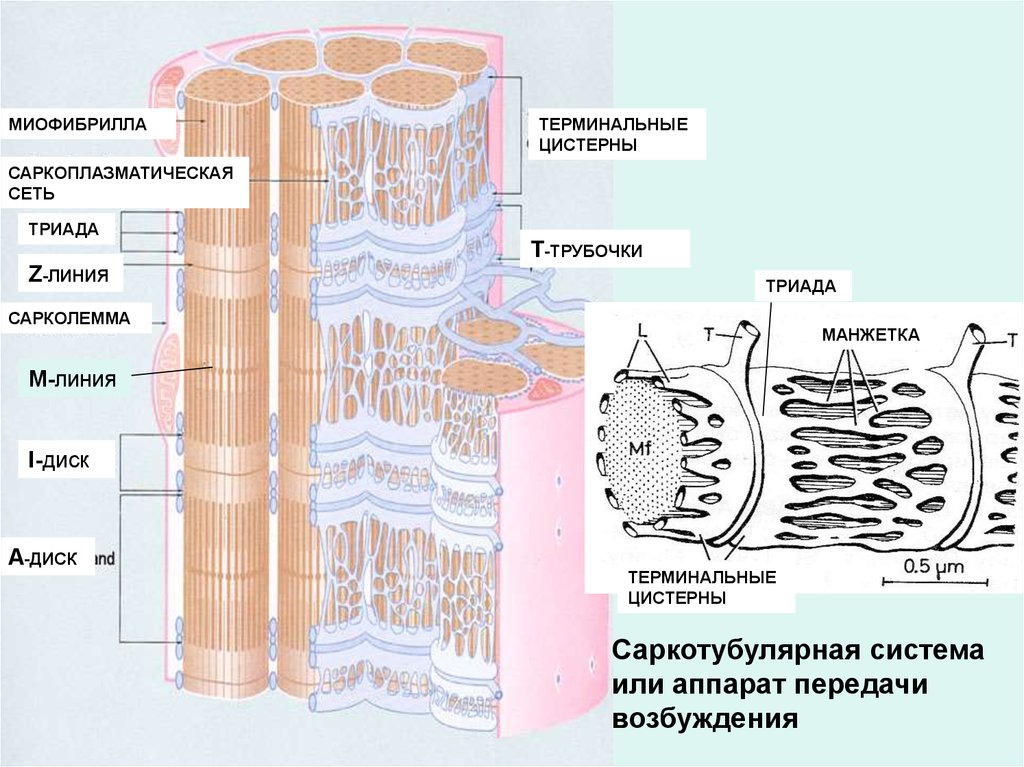
МИОФИБРИЛЛАТЕРМИНАЛЬНЫЕ
ЦИСТЕРНЫ
САРКОПЛАЗМАТИЧЕСКАЯ
СЕТЬ
ТРИАДА
Z-ЛИНИЯ
Т-ТРУБОЧКИ
ТРИАДА
САРКОЛЕММА
МАНЖЕТКА
М-ЛИНИЯ
I-диск
А-ДИСК
ТЕРМИНАЛЬНЫЕ
ЦИСТЕРНЫ
Саркотубулярная система
или аппарат передачи
возбуждения
20.
СХЕМА МЕХАНИЗМОВПЕРЕДАЧИ ВОЗБУЖДЕНИЯ И
ВЫБРОСА КАЛЬЦИЯ ИЗ
САРКОПЛАЗМАТИЧЕСКОЙ
СЕТИ В ПОПЕРЕЧНОПОЛОСАТОЙ МЫШЕЧНОЙ
ТКАНИ
21.
Связывание Ca2+ тонкими нитями. В покое взаимодействие тонких и толстых нитейневозможно, т.к. миозин-связывающие участки F-актина заблокированы тропомиозином. При
высокой концентрации Ca2+ эти ионы связываются с тропонином C и вызывают
конформационные изменения тропомиозина, приводящие к разблокированию миозинсвязывающих участков.
Ca2+–зависимый механизм регуляции взаимодействия актина с миозином В покое
миозин-связывающие участки тонкой нити заняты тропомиозином. При сокращении ионы
Ca2+ связываются с тропонином С, а тропомиозин открывает миозин-связывающие участки.
Головки миозина присоединяются к тонкой нити и вызывают её смещение относительно
толстой нити.
Взаимодействие тонких и толстых нитей. В результате разблокирования миозинсвязывающих участков молекул актина головки миозина, несущие продукты гидролиза АТФ,
присоединяются к тонкой нити и изменяют свою конформацию, создавая тянущее усилие: —
тонкие нити начинают скользить между толстыми. За счёт шарнирного участка в области
шейки миозина происходит гребковое движение, продвигающее тонкую нить к центру
сaркомера. В результате происходит скольжение тонких нитей относительно толстых. Затем
головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от
актина. Последующий гидролиз АТФ восстанавливает конформированную молекулу миозина,
готовую вступить в новый цикл. Такая модель скользящих нитей была предложена Хью
Хаксли.
Головка миозина совершает около пяти циклов в секунду. Когда одни головки миозина
толстой нити производят тянущее усилие, другие в это время свободны и готовы вступить в
очередной цикл. Следующие друг за другом гребковые движения подтягивают тонкие нити к
центру сaркомера. Скользящие тонкие нити тянут за собой Z-линии, вызывая сокращение
сaркомера. Поскольку в процесс сокращения практически одномоментно вовлечены все
саркомеры МВ, происходит его укорочение.
22.
СаЗАВИСИМЫЙ МЕХАНИЗМ РЕГУЛЯЦИИ ВЗАИМОДЕЙСТВИЯ
АКТИНА С МИОЗИНОМ
актин
ТРОПОМИОЗИН
УЧАСТКИ
СВЯЗЫВАНИЯ МИОЗИНА
ТРОПОНИН
АДФ+Ф
23.
ПОСЛЕДОВАТЕЛЬНОСТЬ МЫШЕЧНОГО СОКРАЩЕНИЯ:1. ДЕПОЛЯРИЗАЦИЯ ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЫ И
ГЕНЕРАЦИЯ ПОТЕНЦИАЛА ДЕЙСТВИЯ;
2. РАСПРОСТРАНЕНИЕ ПОТЕНЦИАЛА ДЕЙСТВИЯ ПО
ПЛАЗМОЛЕММЕ МЫШЕЧНОГО ВОЛОКНА;
3. ПЕРЕДАЧА СИГНАЛА В ТРИАДАХ НА САРКОПЛАЗМАТИЧЕСКИЙ
РЕТИКУЛУМ;
4. ВЫБРОС
Са ИЗ САРКОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА;
5. СВЯЗЫВАНИЕ
Са ТРОПРОНИНОМ С ТОНКИХ НИТЕЙ;
6. ВЗАИМОДЕЙСТВИЕ ТОНКИХ И ТОЛСТЫХ НИТЕЙ;
7. ЦИКЛ ВЗАИМОДЕЙСТВИЯ НИТЕЙ;
8. УКОРОЧЕНИЕ САРКОМЕРОВ И СОКРАЩЕНИЕ МЫШЕЧНОГО
ВОЛОКНА;
9. РАССЛАБЛЕНИЕ.
24.
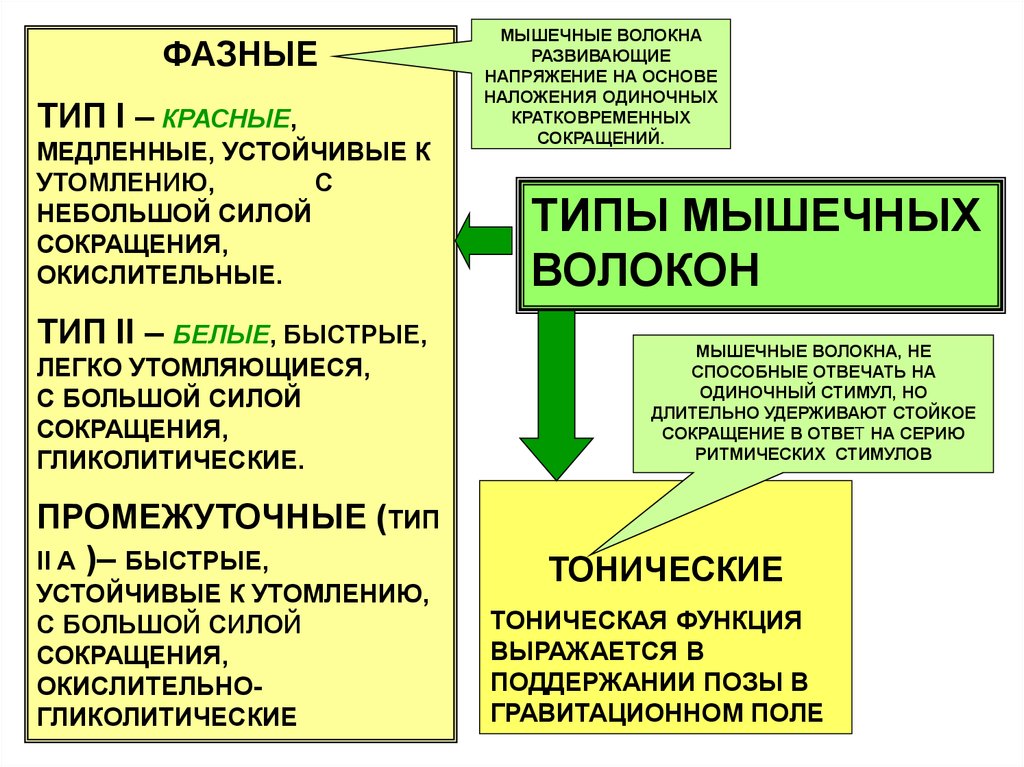
ФАЗНЫЕТИП I – КРАСНЫЕ,
МЕДЛЕННЫЕ, УСТОЙЧИВЫЕ К
УТОМЛЕНИЮ,
С
НЕБОЛЬШОЙ СИЛОЙ
СОКРАЩЕНИЯ,
ОКИСЛИТЕЛЬНЫЕ.
ТИП II – БЕЛЫЕ, БЫСТРЫЕ,
ЛЕГКО УТОМЛЯЮЩИЕСЯ,
С БОЛЬШОЙ СИЛОЙ
СОКРАЩЕНИЯ,
ГЛИКОЛИТИЧЕСКИЕ.
ПРОМЕЖУТОЧНЫЕ (ТИП
II A )– БЫСТРЫЕ,
УСТОЙЧИВЫЕ К УТОМЛЕНИЮ,
С БОЛЬШОЙ СИЛОЙ
СОКРАЩЕНИЯ,
ОКИСЛИТЕЛЬНОГЛИКОЛИТИЧЕСКИЕ
МЫШЕЧНЫЕ ВОЛОКНА
РАЗВИВАЮЩИЕ
НАПРЯЖЕНИЕ НА ОСНОВЕ
НАЛОЖЕНИЯ ОДИНОЧНЫХ
КРАТКОВРЕМЕННЫХ
СОКРАЩЕНИЙ.
ТИПЫ МЫШЕЧНЫХ
ВОЛОКОН
МЫШЕЧНЫЕ ВОЛОКНА, НЕ
СПОСОБНЫЕ ОТВЕЧАТЬ НА
ОДИНОЧНЫЙ СТИМУЛ, НО
ДЛИТЕЛЬНО УДЕРЖИВАЮТ СТОЙКОЕ
СОКРАЩЕНИЕ В ОТВЕТ НА СЕРИЮ
РИТМИЧЕСКИХ СТИМУЛОВ
ТОНИЧЕСКИЕ
ТОНИЧЕСКАЯ ФУНКЦИЯ
ВЫРАЖАЕТСЯ В
ПОДДЕРЖАНИИ ПОЗЫ В
ГРАВИТАЦИОННОМ ПОЛЕ
25.
ТИПЫ МЫШЕЧНЫХ ВОЛОКОН26. РЕГЕНЕРАЦИЯ ПОПЕРЕЧНОПОЛОСАТОЙ МУСКУЛАТУРЫ
27. СЕРДЕЧНАЯ (ИСЧЕРЧЕННАЯ) МЫШЕЧНАЯ ТКАНЬ
28. СТРОЕНИЕ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ
вставочныйдиск
кардтомиоцит
сосуд
кардтомиоцит
вставочный
диск
29.
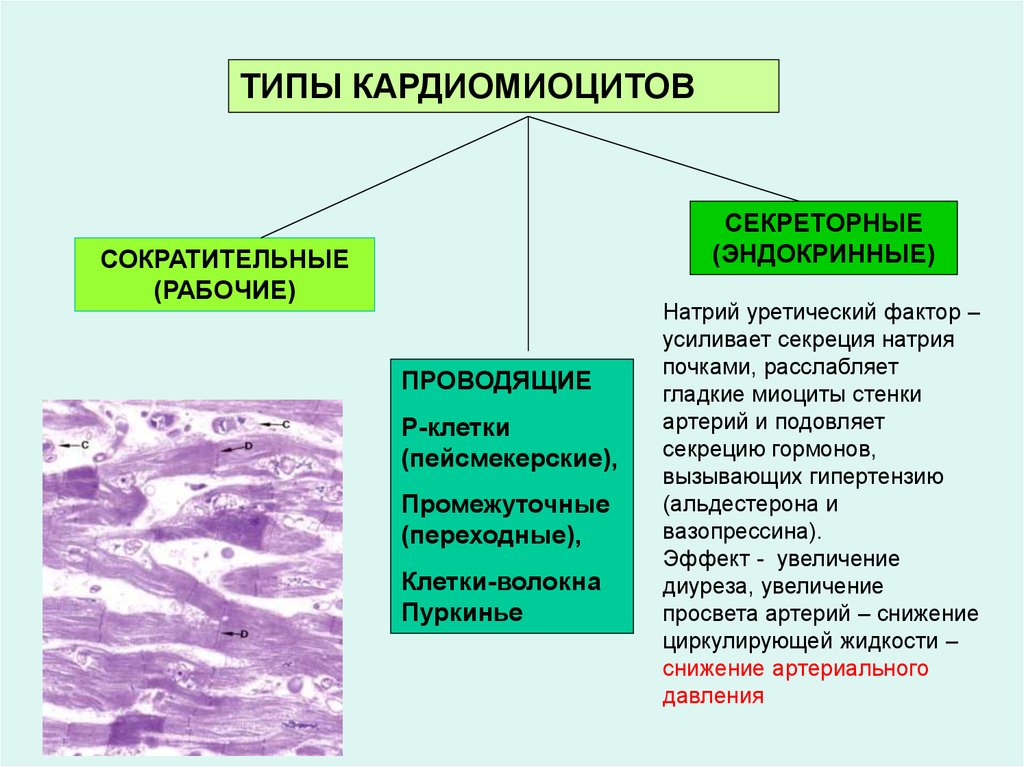
ТИПЫ КАРДИОМИОЦИТОВСЕКРЕТОРНЫЕ
(ЭНДОКРИННЫЕ)
СОКРАТИТЕЛЬНЫЕ
(РАБОЧИЕ)
ПРОВОДЯЩИЕ
Р-клетки
(пейсмекерские),
Промежуточные
(переходные),
Клетки-волокна
Пуркинье
Натрий уретический фактор –
усиливает секреция натрия
почками, расслабляет
гладкие миоциты стенки
артерий и подовляет
секрецию гормонов,
вызывающих гипертензию
(альдестерона и
вазопрессина).
Эффект - увеличение
диуреза, увеличение
просвета артерий – снижение
циркулирующей жидкости –
снижение артериального
давления
30. МИТОХОНДРИАЛЬНАЯ СИСТЕМА КАРДИОМИОЦИТОВ
миофибриллымиофибриллы
межмитохондриальный
контаткт
межмитохондриальный
контаткт
31. САРКОПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ И Т-КАНАЛЫ РАБОЧИХ КАРДИОМИОЦИТОВ
ТТЦ- терминальные цистерны
СР-саркоплазматический
ТЦ
ретикулум
СР
Т-каналы Т-системы
Т
32.
ГЛАДКИЕ МИОЦИТЫ33.
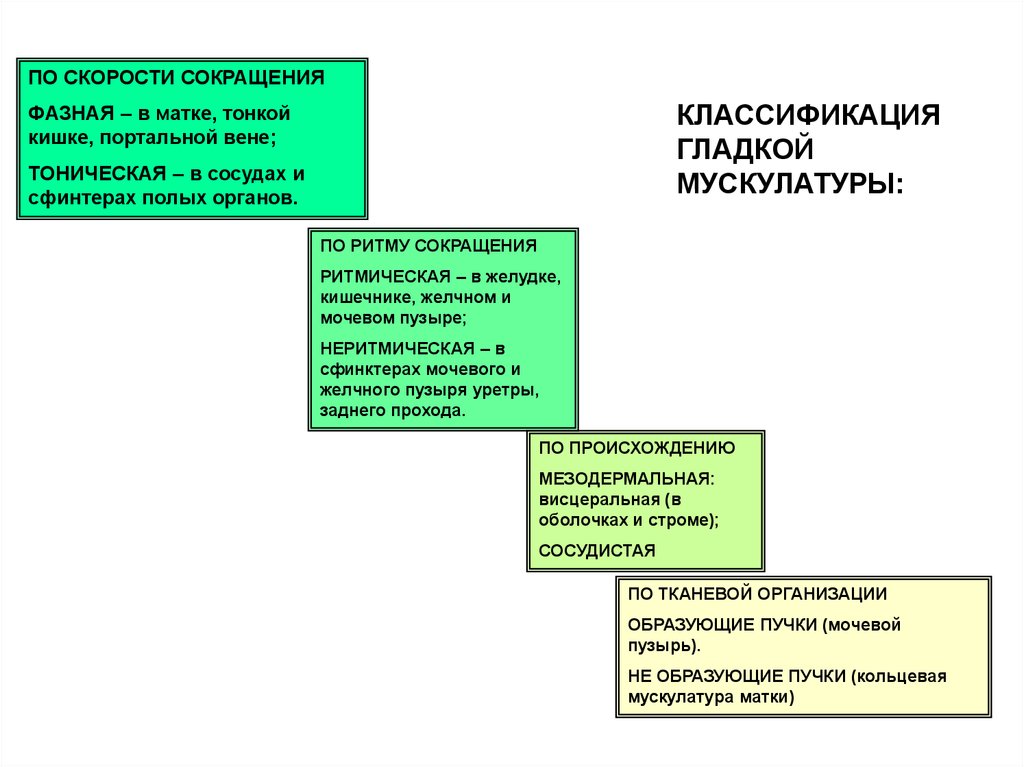
ПО СКОРОСТИ СОКРАЩЕНИЯКЛАССИФИКАЦИЯ
ГЛАДКОЙ
МУСКУЛАТУРЫ:
ФАЗНАЯ – в матке, тонкой
кишке, портальной вене;
ТОНИЧЕСКАЯ – в сосудах и
сфинтерах полых органов.
ПО РИТМУ СОКРАЩЕНИЯ
РИТМИЧЕСКАЯ – в желудке,
кишечнике, желчном и
мочевом пузыре;
НЕРИТМИЧЕСКАЯ – в
сфинктерах мочевого и
желчного пузыря уретры,
заднего прохода.
ПО ПРОИСХОЖДЕНИЮ
МЕЗОДЕРМАЛЬНАЯ:
висцеральная (в
оболочках и строме);
СОСУДИСТАЯ
ПО ТКАНЕВОЙ ОРГАНИЗАЦИИ
ОБРАЗУЮЩИЕ ПУЧКИ (мочевой
пузырь).
НЕ ОБРАЗУЮЩИЕ ПУЧКИ (кольцевая
мускулатура матки)
34.
Различают контрактильные, секреторные и промежуточные леомиоциты.Контрактильные ГМ в них много элементов гладкой ЭПС, слабо развита грЭПС, много
мелких митохондрий, хорошо развит цитоскелет. Основная масса цитоплазмы занята
актиновыми и миозиновыми белками, которые, взаимодействуя друг с другом, образуют
линейные структуры -сократимые единицы. Каждая сократимая единица представляет
собой структуру аналогичную миофибрилле поперечно-полосатого волокна, но в ней нет
правильного распределения тонких и толстых филаментов. Каждая сократимая единица
прикрепляясь в двух точках поверхностного аппарата, под некоторым углом. При
сокращении вся
похожи на фибробласты, но содержат по периферии пучки миофиламентов. По
поверхность клетки приобретает бугристый вид за счет образования глубоких впячиваний
в местах приерепления сократимых единиц.
Секреторные Гм (синтетический фенотип) хорошо развит синтетический аппарат. По
ультраструктуре степени зрелости секреторные леомиоциты относятся к
малодифференцированным. Эти клетки синтезируют протеогликаны, гликопротеиды,
фибронекти, проколлаген и проэластин.
35.
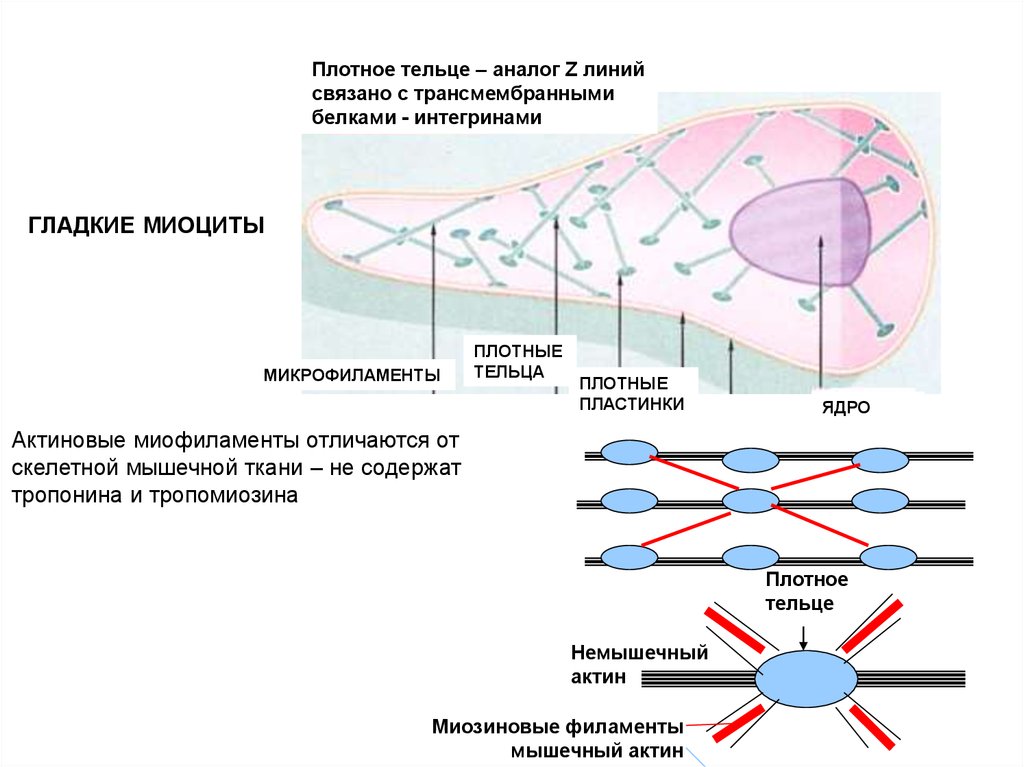
Плотное тельце – аналог Z линийсвязано с трансмембранными
белками - интегринами
ГЛАДКИЕ МИОЦИТЫ
МИКРОФИЛАМЕНТЫ
ПЛОТНЫЕ
ТЕЛЬЦА
ПЛОТНЫЕ
ПЛАСТИНКИ
ЯДРО
Актиновые миофиламенты отличаются от
скелетной мышечной ткани – не содержат
тропонина и тропомиозина
Плотное
тельце
Немышечный
актин
Миозиновые филаменты
мышечный актин
36.
К рождению скелетные мышцы составляют около 25 % массы тела. Самымитолстыми оказываются волокна в диафрагме, самыми тонкими в мышцах голени,
то есть ускоренно формируются мышцы, которые более необходимы для
выполнения жизненно важных функций. Мышцы новорожденного характеризуются
небольшим диаметром волокон, немногочисленностью и поэтому более редким
расположением миофибрилл внутри мышечного волокна. У новорожденного
поперечная исчерченность отчетливее, чем у плода, однако диски I значительно
выше дисков А. К 7—8 годам высота темных и светлых дисков почти одинакова. У
детей разного возраста наблюдаются значительные различия в гистологическом
строении мышц, их кровоснабжении и иннервации. В первые три года количество
миофибрилл в волокнах увеличивается в 4—5 раз, диаметр мышечного волокна —
в 1,5 раза. Изменяется форма ядер: из округлых они становятся палочковидными.
Количество ядер на единицу площади уменьшается, что свидетельствует о росте
волокон в длину. Мышечные волокна из округлых становятся многогранными.
Развиваются кровеносные сосуды, улучшается кровоснабжение мышечных
волокон, активно развивается иннервационный аппарат.
37.
От 3-х до 7 лет количество миофибрилл в волокне увеличивается посравнению с новорожденным в 15—20 раз, соответственно увеличивается
диаметр волокон. Продолжает уменьшаться количество ядер на единицу
площади. В волокнах двуглавой мышцы, например, на единицу площади
приходится у новорожденного 40—45 ядер, у ребенка 10 мес — 28, в 13—
16 лет — 7 ядер. Во всех мышцах разрастается соединительная ткань,
активно растут сухожилия. С 7 до 14 лет рост мышечной ткани резко
ускоряется за счет особенно активного развития количества миофибрилл и
разрастания соединительной ткани. Именно в это время происходит
важнейшая дифференцировка мышцы как органа. Двигательные
нервные окончания приобретают структуру, близкую к таковой у взрослых.
Чувствительная иннервация, имеющая высокий уровень развития у
новорожденного, к 7—8 годам по морфологической организации достигает
уровня, характерного для взрослого. У детей дошкольного возраста сила
мышц незначительна.
38.
Наиболее интенсивно она начинает развиваться в старшем школьномвозрасте между 15 и 18 годами, когда происходит быстрое нарастание массы
мускулатуры. Но нарастание силы мышц опаздывает по сравнению с ростом
мускулатуры на один год. Предполагается, что это связано с возрастными
особенностями развития микроструктуры мышц и организма в целом. С 18 лет
рост силы мышц замедляется (до 26 лет), хотя к этому времени они составляют
уже около 40% от всей массы тела. Развитие мышц, ее сосудистой системы и
иннервации происходит до 25—30 лет.
39.
Хорошо пользоваться скелетной нелокомоторнойи тонической мускулатурой!
Спасибо за внимание!
40.
МЕДИАТОРИЛИ ГОРМОН
Механизмы,
ПОТЕНЦИАЛАКТИВИРУЕМЫЕ
Са КАНАЛЫ
ХЕМОАКТИВИРУЕМЫЕ
Са КАНАЛЫ
повышающие
БЕЛОКРЕЦЕПТОР
Ca²
уровень Са
Са КАНАЛЫ,
ОТКРЫВАЕМЫЕ
ДЕПОЛЯРИЗАЦИЕЙ
в гладких
миоцитах
А
Ca²
Ca²
Фосфолипаза С
Ca²
Б
Са АТФАЗА
МЕДИАТОР
ИЛИ ГОРМОН
БЕЛОК-РЕЦЕПТОР,
СЦЕПЛЕННЫЙ С
ФОСФОЛИПАЗОЙ
ВЫБРОС Са
ИЗ РЕТИКУЛУМА
В
УДАЛЕНИЕ Са²
Г