










































 biology
biologySimilar presentations:










Биологические процессы развития зародыша. Ткани. Эпителиальная ткань. (Лекция 2)
1.
Лекция №22.
Лекция №2Представление о биологических процессах, лежащих
в основе развития зародыша - индукция,
детерминация, деление, миграция клеток, рост,
дифференцировка,
взаимодействие
клеток,
разрушение.
Ткани. Понятие о диффероне или гистогенетическом
ряде. Регенерация тканей.
Эпителиальные ткани. Основные типы покровного
и железистого эпителия. Источники развития.
Железы
3.
• Причи́на:• 1) основание, предлог для каких-нибудь действий
• Пример: Уважительная причина; Смеяться без
причины; По причине того что..., по той причине что...,
из-за того что...2) явление, вызывающее,
обусловливающее возникновение другого явления
• Пример: Причина пожара; Причина спешки в том, что не
хватает времени. Ещё неоплатонический философ
Прокл (в своём комментарии к диалогу Платона Тимею)
насчитывает у одного Платона 64 различных понятия о
причине, а у Аристотеля — 48. Это число можно
сократить до двух основных понятий причины у Платона и
до четырёх — у Аристотеля.
4.
эмбриональная индукция — взаимодействие элементовразвивающегося зародыша, при котором воздействие
одного из них направляет (индуцирует) развитие другого.
NB! В результате такого взаимодействия запускается
цепь морфогенетических (формообразовательных)
процессов. Элемент, оказывающий воздействие,
назван индуктором. Способность воспринимать
индукционное воздействие и отвечать на него адекватным
образом определяется как компетенция, а элемент
(структура, клеточная группа) организма, способный
реагировать на индукционное воздействие изменением
своего развития, назван компетентной тканью.
5.
В результате компетентная ткань становится детерминированной (предопределенной) к специфическому типуразвития (к развитию с определенным результатом).
Детерминированное состояние (определенный результат
развития) реализуется в виде дифференциации структур
и/или частей (фрагментов) развивающегося организма, а
также клеточной дифференцировки.
Нередко (может быть и всегда) основу индукционных
взаимоотношений составляют межклеточные
взаимодействия, без которых не обходится ни один акт
развития.
6.
Феноменэмбриональной
индукции
был открыт
немецким эмбриологом
Региональная
специфичность
индукции:
трансплантация
различных
Г. Шпеманом
и крыши
его ученицей
Г. Мангольд
в 1921 гзародыша
(выращивание
химер)
участков
гастроцеля
в бластоцель
(амфибии)
Ранняя нейрула
Ранняя гаструла
Трансплантация
части крыши
гастроцеля
7. ДЕТЕРМИНИРОВАННЫЙ и РЕГУЛЯТИВНЫЙ типы РАЗВИТИЯ ЖИВОТНЫХ -
ДЕТЕРМИНИРОВАННЫЙ и РЕГУЛЯТИВНЫЙтипы РАЗВИТИЯ ЖИВОТНЫХ ДЕТЕРМИНАЦИЯ АКТОВ РАЗВИТИЯ (по результату)
ЯВЛЯЕТСЯ НЕПРЕМЕННОЙ ХАРАКТЕРИСТИКОЙ, но в различных
систематических группах ЖИВОТНЫХ ПРЕДСТАВЛЕНА с
ОСОБЕННОСТЯМИ: ВЫДЕЛЯЮТ ДЕТЕРМИНИРОВАННЫЙ и
РЕГУЛЯТИВНЫЙ типы РАЗВИТИЯ;
ДЕТЕРМИНИРОВАННЫЙ (МОЗАИЧНЫЙ) тип – СУДЬБЫ
КЛЕТОК в РАЗВИТИИ определяются РАНО (БЛАСТОМЕРЫ);
ТИПИЧЕН, в основном, для ПЕРВИЧНОРОТЫХ (ГРЕБНЕВИКИ, КРУГЛЫЕ и
КОЛЬЧАТЫЕ ЧЕРВИ, МОЛЛЮСКИ) и некоторых ВТОРИЧНОРОТЫХ
(АСЦИДИИ);
РЕГУЛЯТИВНЫЙ тип - СУДЬБЫ КЛЕТОК в РАЗВИТИИ
определяются ПОЗЖЕ (БЛАСТУЛА, ГАСТРУЛА); ТИПИЧЕН для
ВТОРИЧНОРОТЫХ, в частности, почти для всех ХОРДОВЫХ;
8. От чего зависит тип развития?
тип РАЗВИТИЯ зависит от того, как РАНО ПРОИСХОДИТСМЕНА СВОЙСТВ КЛЕТОК:
ТОТИПОТЕНТНОСТЬ → МУЛЬТИПОТЕНТНОСТЬ
(ЛАБИЛЬНАЯ ДЕТЕРМИНАЦИЯ, КОММИТАЦИЯ) →
УНИПОТЕНТНОСТЬ (СТАБИЛЬНАЯ или
ОКОНЧАТЕЛЬНАЯ ДЕТЕРМИНАЦИЯ, КОММИТАЦИЯ);
СУДЬБА ЭЛЕМЕНТА СИСТЕМЫ (КЛЕТКИ) в
РАЗВИТИИ есть ФУНКЦИЯ его ПОЛОЖЕНИЯ в ЦЕЛОМ
(место среди всех клеток, место в «иерархии» клеток и
т.п.).
9.
КОНЦЕПЦИЯ “ПОЗИЦИОННОЙ ИНФОРМАЦИИ” –ПЕРИОДИЧЕСКИ КЛЕТКА ПОЛУЧАЕТ “ДОЗУ Позиционной
Информации”, чем и ОПРЕДЕЛЯЕТСЯ её ПОВЕДЕНИЕ на
очередном ЭТАПЕ РАЗВИТИЯ;
КОНЦЕПЦИЯ “МОРФОГЕНЕТИЧЕСКОГО ПОЛЯ” - ОКАЗАВШИСЬ в
ОПРЕДЕЛЕННОМ РЕГИОНЕ в ТЕЛЕ ЗАРОДЫША (МОРФОГЕНЕТИЧЕСКИЙ КОМПАРТМЕНТ) КЛЕТКА ИДЕНТИФИЦИРУЕТ УСЛОВИЯ и
КООРДИНИРУЕТ с ними своё ПОВЕДЕНИЕ;
в обоих случаях КЛЕТКА ОПРЕДЕЛЯЕТСЯ с
ПОВЕДЕНИЕМ: ПРОЛИФЕРАЦИЯ на месте или
МИГРАЦИЯ, ДИФФЕРЕНЦИРОВКА или
АПОПТОЗ, АССОЦИАЦИЯ с другими КЛЕТКАМИ, что в своей
основе имеет СМЕНУ АКТИВНЫХ ГЕНОВ КЛЕТОЧНОГО
ГЕНОМА;
10. КАСКАД АКТОВ ЭМБРИОНАЛЬНОЙ ИНДУКЦИИ –
МЕЗОДЕРМАЛИЗУЮЩИЙ АКТ ИНДУКЦИИ – ИСТИННАЯПЕРВИЧНАЯ (НЬЮКУПОВСКАЯ) ЭМБРИОНАЛЬНАЯ ИНДУКЦИЯ: в
роли ИНДУКТОРА ВЫСТУПАЕТ ЭНТОДЕРМА, которая ПОБУЖДАЕТ
определённую КЛЕТОЧНУЮ ГРУППУ к РАЗВИТИЮ в НАПРАВЛЕНИИ
МЕЗОДЕРМЫ – ДОРЗАЛЬНОЙ и ВЕНТРАЛЬНОЙ; ОРГАНИЗУЕТ ДОРЗОВЕНТРАЛЬНУЮ РЕГИОНАЛИЗАЦИЮ и ЛЕВО-ПРАВУЮ АССИМЕТРИЮ
ПРОЦЕССОВ РАЗВИТИЯ;
НЕЙРАЛИЗУЮЩИЙ АКТ ИНДУКЦИИ – КЛАССИЧЕСКАЯ
ПЕРВИЧНАЯ (ШПЕМАНОВСКАЯ) ЭМБРИОНАЛЬНАЯ ИНДУКЦИЯ: в
роли ИНДУКТОРА ВЫСТУПАЕТ ДОРЗАЛЬНАЯ ГУБА БЛАСТОПОРА
(ХОРДОМЕЗОДЕРМАЛЬНЫЙ ЗАЧАТОК), который ПОБУЖДАЕТ
определённый РЕГИОН ЭКТОДЕРМЫ к РАЗВИТИЮ в НАПРАВЛЕНИИ
НЕРВНОЙ ТРУБКИ; ОРГАНИЗУЕТ РОСТРО-КАУДАЛЬНУЮ
РЕГИОНАЛИЗАЦИЮ ПРОЦЕССОВ РАЗВИТИЯ;
СТРУКТУРО/ОРГАНОСПЕЦИФИЧНЫЕ АКТЫ ИНДУКЦИИ –
ЧАСТНЫЕ СТРУКТУРО/ОРГАНОГЕНЕЗЫ в пределах ОРГАННЫХ
МОРФОГЕНЕТИЧЕСКИХ ПОЛЕЙ; ЭПИТЕЛИО-МЕЗЕНХИМАЛЬНАЯ
ИНДУКЦИЯ;
NB! Гистогенез – органогенез
(формирование «общего», а затем «частного»)
Понятие «ТКАНЬ»
11.
ТКАНИ:ИСТОРИЧЕСКИ СЛОЖИВШАЯСЯ СИСТЕМА ЭЛЕМЕНТОВ (КЛЕТОК И НЕКЛЕТОЧНОГО
ЖИВОГО ВЕЩЕСТВА), ОБЪЕДИНЕННЫХ МЕЖДУ СОБОЙ ОБЩЕЙ СТРУКТУРОЙ, ФУНКЦИЕЙ И РАЗВИТИЕМ…….(А.А.Заварзин)
СИСТЕМА КЛЕТОК И НЕКЛЕТОЧНЫХ СТРУКТУР, ИМЕЮЩИХ ОБЩЕЕ СТРОЕНИЕ,
РАЗВИТИЕ И ФУНКЦИИ (М.Я.Субботин)
СИСТЕМА ВЗАИМОДЕЙСТВУЮЩИХ КЛЕТОЧНЫХ ДИФФЕРОНОВ, ВОЗНИКНОВЕНИЕ,
СТРОЕНИЕ И ФУНКЦИИ КОТОРЫХ ДЕТЕРМИНИРОВАНЫ ИСТОРИЧЕСКИМ И ИНДИВИДУАЛЬНЫМ РАЗВИТИЕМ (Р.К.Данилов)
ДИФФЕРОН – РЯД ПОСЛЕДОВАТЕЛЬНО ДИФФЕРЕНЦИРУЮЩИХСЯ
КЛЕТОК ОТ МАЛОДИФФЕРЕНЦИРОВАННЫХ НЕЗРЕЛЫХ ДО ВЫСОКОДИФФЕРЕНЦИРОВАННЫХ СПЕЦИАЛИЗИРОВАННЫХ ФОРМ.
12.
В образовании ткани могут принимать участие следующиеэлементы:
•Клетки (производные клеток - симпласты, синцитии,
постклеточные структуры - эритроциты и тромбоциты)
•Аморфное межклеточное вещество
•Волокна (коллагеновые, эластические, ретикулярные)
100
80
60
клетки
40
волокна
ам. в-во
20
0
эпителий
кровь
РВСТ
ПВСТ
Хрящ
кость
мышцы
Каждая ткань отличается определённым составом таких элементов.
Например, скелетная мышечная ткань - это лишь симпласты
(мышечные волокна), кровь - набор определённых клеток в
определённой межклеточной среде и т.д. Этот состав
(соотношение) и физические/химические/механические свойства
элементов ткани обуславливает специфические функции каждой
ткани. Выполняя эти функции, элементы тканей обычно тесно
взаимодействуют между собой, образуя единое целое.
13.
Этапы развития тканей в онтогенезе:• этап топической дифференцировки —предположительные зачатки тканей оказываются в определённых зонах
цитоплазмы яйцеклетки, а затем и зиготы;
• этап бластомерной дифференцировки — в результате
дробления зиготы зачатки тканей оказываются
локализованными в разных бластомерах зародыша;
• этап зачатковой дифференцировки — в результате
гаструляции зачатки тканей локализованы в различных
участках зародышевых листков;
• гистогенез — процесс преобразования зачатков тканей в
ткани в результате пролиферации, роста, индукции,
детерминации, миграции и дифференцировки клеток.
14.
Основные теории развития тканей в филогенезе:• Закон параллельных рядов (А. А. Заварзин) — ткани
животных разных классов и видов, выполняющие
одинаковые функции, имеют сходное строение, так
как развиваются они параллельно у разных животных
филогенетического древа; примеры – все ткани
• Закон дивергентной эволюции тканей (Н. Г. Хлопин)
— в филогенезе происходит расхождение признаков
тканей и появление новых разновидностей ткани в
пределах тканевой группы, что приводит к
усложнению животных организмов и увеличению
разнообразия тканей. Примеры – строение эпидермиса
и его производных морских животных, земноводных,
птиц и млекопитающих
15.
Дифференцировка – стойкоеструктурно-функциональное изменение
ранее однородных клеток.
Определяется детерминацией, или
закодированной в ДНК программой
дифференцировки клеток.
В процессе дифференцировки
формируется пул (популяция) активно
функционирующих клеток.
16.
ВременнАя дифференцировка – этапностьизменений клеток в составе ткани.
СЛОИ
роговой
блестящий
зернистый
шиповатый
базальный
СОЕДИНИТЕЛЬНАЯ ТКАНЬ
17.
Пространственная дифференцировка –образование различных типов
специализированных клеток в составе ткани.
В СОСТАВ РЕСНИТЧАТОГО
ЭПИТЕЛИЯ ВХОДЯТ:
РЕСНИТЧАТЫЕ КЛЕТКИ
БОКАЛОВИДНЫЕ КЛЕТКИ
БАЗАЛЬНЫЕ КЛЕТКИ
ПРОМЕЖУТОЧНЫЕ
(ВСТАВОЧНЫЕ) КЛЕТКИ
ЭНДОКРИННЫЕ КЛЕТКИ
18.
Биохимическая дифференцировка –образование клеток ткани, секретирующих
специфические типы белков.
ЭКЗОКРИННЫЕ И ЭНДОКРИННЫЕ КЛЕТКИ КИШЕЧНОГО ЭПИТЕЛИЯ
19.
в первую очередь дифференцируются стволовыеклетки, дающие начало дифферону.
Признаки стволовых клеток:
1. Способность к самоподдержанию популяции.
2. Способность к делению. NB! Предел Хейфлика.
3. Способность части клеток дифференцироваться
после деления. NB! Отсюда - два вида делений!
Процесс дифференцировки клеток тканей регулируется нервной, эндокринной и иммунной системами и
тканевыми механизмами регуляции.
Кейлоны – вещества, вырабатываемые
дифференцированными клетками и способные
тормозить пролиферацию и дифференцировку
стволовых клеток.
Коммитирование – ограничение путей развития.
Пример – бластомеры зиготы.
20.
Предел или лимит Хейфлика (Hayflick limit) — границаколичества делений соматических клеток. В 1961 году
Хейфлик наблюдал, как фибробласты человека, делящиеся
в клеточной культуре, умирают приблизительно после 50
делений и проявляют признаки старения при приближении к
этой границе. Эта граница была найдена в культурах всех
полностью дифференцированных клеток как человека, так и
других многоклеточных организмов. (NB! Объяснить
эксперимент Хейфлика)
Максимальное число делений клетки различно в
зависимости от типа клетки и ещё сильнее различается в
зависимости от организма, которому эта клетка
принадлежит. Для большинства человеческих
клеток предел Хейфлика составляет 52
деления.
21.
Граница Хейфлика связана с сокращениемразмера теломер, участков ДНК на концах хромосом.
Теломеры укорачиваются медленно — по несколько (3-6)
нуклеотидов за клеточный цикл, то есть за 52 цикла они
укоротятся всего на 150—300 нуклеотидов. Вывод- чем
короче у ДНК теломеры, тем больше делений у нее прошло,
а значит - тем старше клетка.
Фермент теломераза – способствует удлинение теломер,
при этом удлиняется и жизнь клетки. Клетки, в которых
функционирует теломераза (половые, раковые),
бессмертны. В соматических клетках, из которых в основном
и состоит организм, теломераза "не работает", поэтому
теломеры при каждом делении клетки укорачиваются, что в
конечном итоге приводит к ее гибели в пределах лимита
Хейфлика, потому что другой фермент - ДНК-полимераза не
способен реплицировать концы молекулы ДНК.
22.
В настоящее время предложена эпигенетическая теория старения,которая объясняет эрозию теломер прежде всего активностью клеточных
рекомбиназ, активизирующихся в ответ на повреждения ДНК,
вызванные, главным образом, возрастной депрессией мобильных
элементов генома. Когда после определённого числа делений теломеры
исчезают совсем, клетка замирает в определённой стадии клеточного
цикла или запускает программу апоптоза.
23.
Организменный уровень интеграцииТеория изнашивания - Sacher, 1966
Теория катастрофы ошибок - Orgel, 1963
Теория стрессового повреждения - Stlye, 1970
Теория аутоинтоксикации - Metchnikoff, 1904
Эволюционная теория (теория программированного старения) Williams, 1957
Теория сохранения информации (теория программированного
старения)
Органный уровень
Эндокринная теория - Korenchevsky, 1961
Иммунологическая теория - Walford, 1969
Торможение головного мозга
Клеточный уровень
Теория клеточных мембран - Zg-Nagy, 1978
Теория соматических мутаций - Szillard, 1959
Митохондриальная теория - Miquel et al., 1980
Митохондриально-лизосомальная теория - Brunk, Terman, 2002
Теория пролиферативного лимита клетки (теория
программированного старения) - Hayflick, Moorhead, 1961
24.
Молекулярный уровеньТеория накопление повреждений ДНК - Vilenchik, 1970
Теория следовых элементов - Eichhorn, 1979
Свободно-радикальная теория - Harman, 1956
Теория поперченных сшивок - Bjorksten, 1968
Теория окислительного стресса - Sohal, Allen, 1990; Yu, Yang, 1996
Теория неэнзиматической гликозиляции - Cerami, 1985
Теория карбонильной интоксикации - Yin, Brunk, 1995
Теория катастрофы загрязнения - Terman, 2001
Теория генных мутаций
Теория укорочения теломер (теория программированного старения) Оловников, 1971
Прочие подходы
Cтарение как энтропия - Sacher, 1967; Bortz, 1986
Математические теории и различные унифицированные теории Sohal, Alle, 1990; Zg-Nagy, 1991; Kowald, Kirkwood, 1994
25.
Коммитирование - постепенное ограничениевозможных направлений развития клеток. Во
взрослом организме - при дифференцировке
полипотентных стволовых клеток крови (на
определённой стадии дифференцировки
превращаются в 8 видов унипотентных клеток,
каждая из которых может развиваться только в один
вид клеточных элементов крови).
Механизм коммитирования - стойкая репрессия
одних и дерепрессия других генов (в клетках
постепенно меняется спектр фунционально
активных генов, что определяет всё более узкое и
конкретное направление дальнейшего развития
клеток).
26.
Детерминация - на определённой стадии коммитирование приводит к тому, что у клетки остаётся толькоодин путь развития - то есть появление у клетки
генетической запрограммированности только на
один путь развития.
Таким образом, детерминация - более узкое понятие,
чем коммитирование: превращение тотипотентных
клеток в полипотентные, олигопотентные и, наконец,
унипотентные - это всё коммитирование;
детерминация - на самом последнем этапе при
образовании унипотентных клеток.
27.
Обеспечение гомеостазаСтационарное состояние дифферонов - когда в
диффероне постоянно происходит процесс
дифференцировки (например, в эпидермисе),
устанавливается стационарное состояние:
каждая клеточная форма дифферона образуется с
такой же скоростью, с какой происходит её убыль
(в результате перехода в последующие формы,
отмирания или удаления).
28.
Ещё раз !!!: два типа делений или два типа потомковстволовых клеток
Для поддержания такого состояния необходимо, чтобы
стволовые клетки не только регулярно вступали в
дифференцировку, но и постоянно пополняли свой запас.
Это обеспечивается за счёт двух типов деления стволовых
клеток - "дифференцировочных": дочерние клетки
вступают в процесс дифференцировки; и
"недифференцировочных": дочерние клетки сохраняют все
свойства стволовых клеток.
Такая способность обозначается, как способность к
самоподдержанию популяции. Это одно из ключевых
свойств стволовых клеток..
29.
Регуляция кейлонами - дифференцировка находится подгуморальным контролем.
Один из способов такого контроля - отрицательная
обратная связь.
Дифференцированные клетки выделяют кейлоны ингибиторы клеточных делений. Когда зрелых клеток
много, под действием их кейлонов деления
предшествующих клеток происходят редко; при недостатке
зрелых клеток ослабевает кейлоновое торможение, и в
созревание вступает большее количество стволовых клеток.
30.
Другие способы регуляции - в эмбриональном периоде действие тканевых индукторов.Например, хорда выделяет индукторы развития нервной
трубки.
После рождения на некоторые виды дифференцировки
влияют гормоноподобные вещества: почки синтезируют
эритропоэтин, стимулирующий эритропоэз в красном
костном мозге. При недостатке эритроцитов в крови
(например, в результате кровопотери) выработка
эритропоэтина усиливается.
Это свидетельствует о наличии отрицательной обратной
связи: зрелые клеточные формы (эритроциты) при
достаточной концентрации в крови тормозят выработку
индукторов своего образования.
31.
Регенерация – способность тканей к восстановлению послеестественной гибели или повреждения. В различных тканях регенерация
протекает неодинаково.
Внутриклеточная регенерация – восстановление
органелл. Характерна для клеток нервной ткани, миокарда, слюнных
желез и клеток печени, так как в этих органах нет стволовых клеток.
Обеспечивает жизнедеятельность клеток на заданном уровне.
Клеточная регенерация – осуществляется за счет деления
клеток. Характерна для тканей, в которых присутствуют стволовые клетки
(эпителий, скелетная мышечная ткань и проч.).
Гистотипическая регенерация – замещение специфических
структур органа (клеток паренхимы) соединительной тканью (стромой).
32.
Физиологическая регенерация – восстановление клетоктканей после их естественной гибели.
Репаративная регенерация – восстановление клеток ткани
или органа после их повреждения.
33.
1665 год. Роберт Гук при помощи примитивного микроскопасобственной конструкции увидел ячейки в тонком срезе коры
пробкового дерева.
1830 год. Ян Пуркинье обнаружил в клетке цитоплазму.
1833 год. Броун обнаружил в клетке ядро.
1838 год. Мюллер и Шванн на основе вывода, сходстве строения
клеток разных организмов сформулировали клеточную теорию
1858 год. Вирхов установил, что новые клетки образуются в
результате деления материнской клетки.
1866 год. Р. Кёлликер классифицировал ткани на
4 вида.
1934 год. А.А. Заварзин установил закон параллельных рядов в
эволюции тканей.
34.
ТКАНИ1.ЭПИТЕЛИАЛЬНЫЕ
2.ОПОРНО-ТРОФИЧЕСКИЕ (СОЕДИНИТЕЛЬНЫЕ)
3.МЫШЕЧНЫЕ
4.НЕРВНАЯ
35.
Эпителии–
пограничные
ткани
организма,
образующие покров тела, слизистые оболочки,
железы.
Источниками эмбрионального развития эпителиев
являются эктодерма, энтодерма, промежуточная и
латеральные (спланхнотом) части мезодермы, а
также мезенхима (эндотелий сосудов, камер сердца).
NB! Отсутствует единый источник происхожения
Практическая значимость гистологии эпителиев
связана с их наличием во всех органах. Такие
патологические образования как полипы, аденомы,
раковые опухоли развиваются из эпителиев
36.
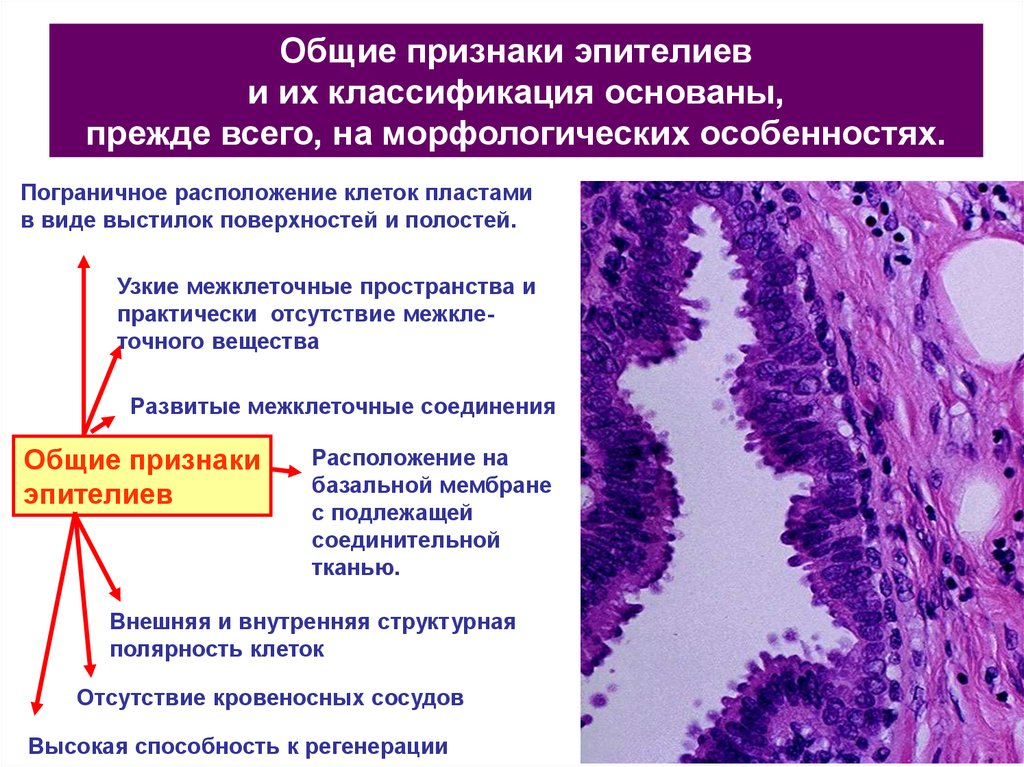
Общие признаки эпителиеви их классификация основаны,
прежде всего, на морфологических особенностях.
Пограничное расположение клеток пластами
в виде выстилок поверхностей и полостей.
Узкие межклеточные пространства и
практически отсутствие межклеточного вещества
Развитые межклеточные соединения
Общие признаки
эпителиев
Расположение на
базальной мембране
с подлежащей
соединительной
тканью.
Внешняя и внутренняя структурная
полярность клеток
Отсутствие кровеносных сосудов
Высокая способность к регенерации
37.
ПО ФУНКЦИИ ЭПИТЕЛИИКЛАССИФИЦИРУЮТСЯ НА
ПОКРОВНЫЕ
ЖЕЛЕЗИСТЫЕ
СМЕШАННЫЕ
38.
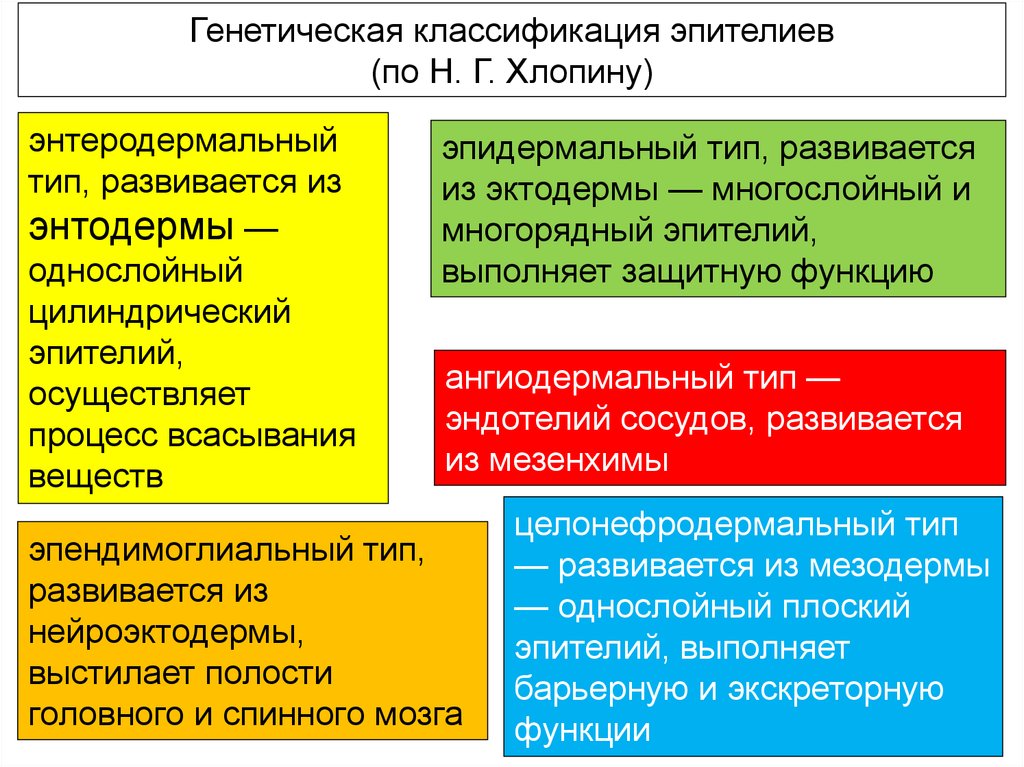
Генетическая классификация эпителиев(по Н. Г. Хлопину)
энтеродермальный
тип, развивается из
энтодермы —
однослойный
цилиндрический
эпителий,
осуществляет
процесс всасывания
веществ
эпидермальный тип, развивается
из эктодермы — многослойный и
многорядный эпителий,
выполняет защитную функцию
ангиодермальный тип —
эндотелий сосудов, развивается
из мезенхимы
эпендимоглиальный тип,
развивается из
нейроэктодермы,
выстилает полости
головного и спинного мозга
целонефродермальный тип
— развивается из мезодермы
— однослойный плоский
эпителий, выполняет
барьерную и экскреторную
функции
39.
Полярность эпителиальных клеток, особенно выраженная воднослойных эпителиях, проявляется в специализации
поверхности (производные плазмолеммы) и характерном
расположении органелл, особенно выраженном в секреторных
клетках.
Производные
плазмолеммы
Апикальные
1. Микроворсинки,
щеточные и
исчерченные каемки,
стереоцилии.
2. Реснички
Латеральные
Межклеточные контакты
Базальные
1.Полудесмосомы.
2. Базальная исчерченность.
40.
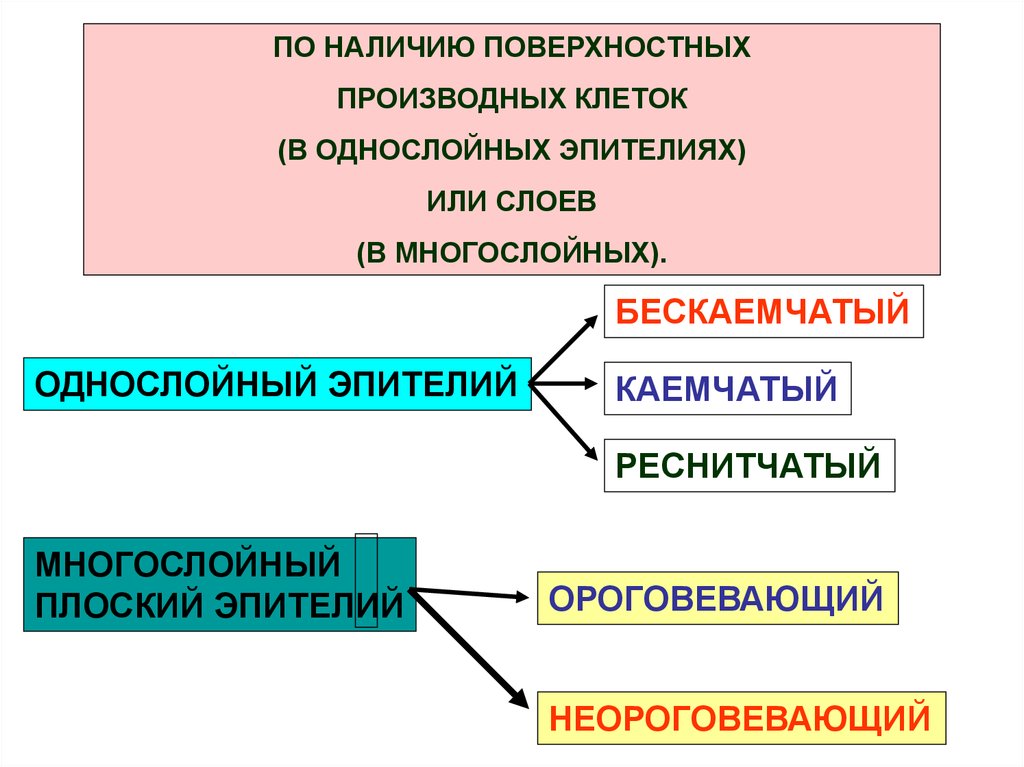
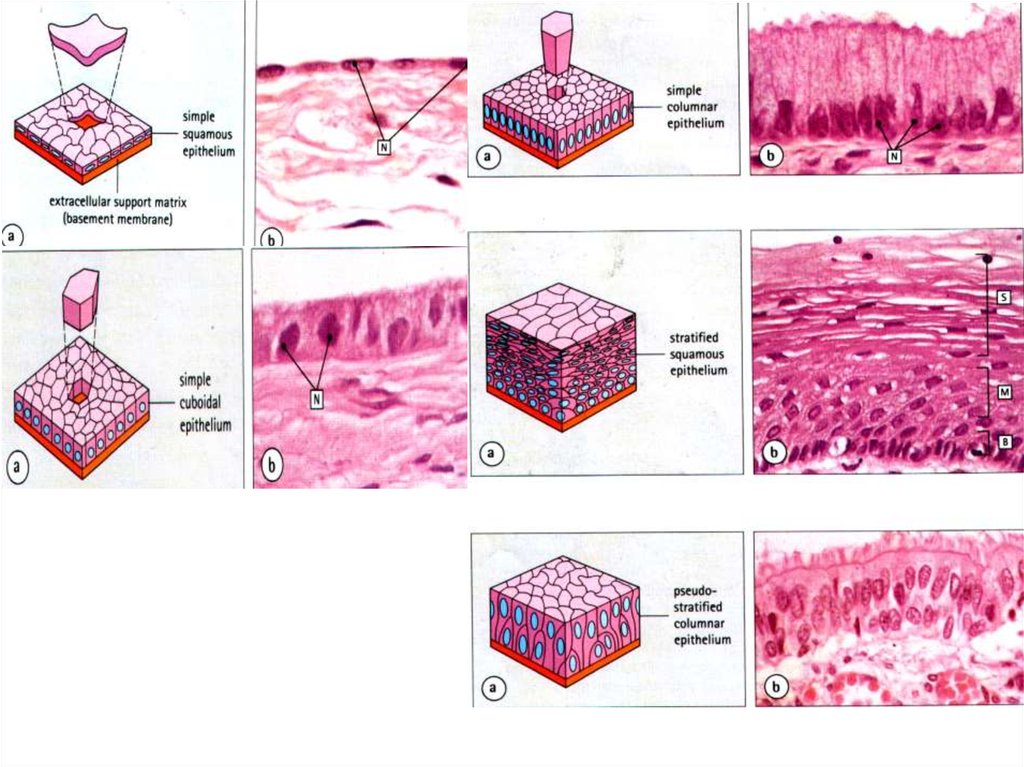
МОРФОЛОГИЧЕСКАЯКЛАССИФИКАЦИЯ ЭПИТЕЛИЕВ
ПО ОТНОШЕНИЮ
КЛЕТОК К БАЗАЛЬНОЙ
МЕМБРАНЕ.
ПО ФОРМЕ КЛЕТОК
ПО НАЛИЧИЮ
ПОВЕРХНОСТНЫХ
ПРОИЗВОДНЫХ КЛЕТОК
(В ОДНОСЛОЙНЫХ ЭПИТЕЛИЯХ)
ИЛИ СЛОЕВ
(В МНОГОСЛОЙНЫХ)
41.
ПО ОТНОШЕНИЮ КЛЕТОКК БАЗАЛЬНОЙ МЕМБРАНЕ
ВСЕ КЛЕТКИ СОЕДИНЕНЫ
С БАЗАЛЬНОЙ МЕМБРАНОЙ
С БАЗАЛЬНОЙ МЕМБРАНОЙ
СОЕДИНЕНЫ ТОЛЬКО КЛЕТКИ
САМОГО ГЛУБОКОГО (БАЗАЛЬНОГО)
СЛОЯ. ОСТАЛЬНЫЕ – ДРУГ С ДРУГОМ, С
ВЫШЕ- И НИЖЕЛЕЖАЩИМИ.
ОДНОСЛОЙНЫЕ
ОДНОСЛОЙНЫЕ МНОГОРЯДНЫЕ
(ПСЕВДОМНОГОСЛОЙНЫЕ)
МНОГОСЛОЙНЫЕ
ПЕРЕХОДНЫЙ
42.
ПО ФОРМЕ КЛЕТОКПЛОСКИЕ
ПРИЗМАТИЧЕСКИЕ
КУБИЧЕСКИЕ
ДЛЯ МНОГОСЛОЙНЫХ ЭПИТЕЛИЕВ КРИТЕРИЕМ
ЯВЛЯЕТСЯ ФОРМА КЛЕТОК ПОВЕРХНОСТНОГО СЛОЯ
43.
ПО НАЛИЧИЮ ПОВЕРХНОСТНЫХПРОИЗВОДНЫХ КЛЕТОК
(В ОДНОСЛОЙНЫХ ЭПИТЕЛИЯХ)
ИЛИ СЛОЕВ
(В МНОГОСЛОЙНЫХ).
БЕСКАЕМЧАТЫЙ
ОДНОСЛОЙНЫЙ ЭПИТЕЛИЙ
КАЕМЧАТЫЙ
РЕСНИТЧАТЫЙ
МНОГОСЛОЙНЫЙ
ПЛОСКИЙ ЭПИТЕЛИЙ
ОРОГОВЕВАЮЩИЙ
НЕОРОГОВЕВАЮЩИЙ
44.
45.
СЕКРЕТОРНЫЕ ЭПИТЕЛИИ И ЖЕЛЕЗЫЭКЗОКРИННЫЕ ЖЕЛЕЗЫ
ЭКЗОКРИННЫЕ
ЭНДОКРИННЫЕ
ПРОСТЫЕ (проток неразветвлен)
СЛОЖНЫЕ (проток разветвлен)
АЛЬВЕОЛЯРНЫЕ
ТРУБЧАТЫЕ
СМЕШАННЫЕ
ВЫРАБАТЫВАЮЩИЕ
БЕЛКОВЫЕ СЕКРЕТЫ
ВЫРАБАТЫВАЮЩИЕ
СЛИЗИСТЫЕ СЕКРЕТЫ
ВЫРАБАТЫВАЮЩИЕ
ПРОИЗВОДНЫЕ ЛИПИДОВ
СЕКРЕТИРУЮЩИЕ
ИОНЫ
РАЗВЕТВЛЕННЫЕ (концевой отдел
разветвлен)
НЕРАЗВЕТВЛЕННЫЕ (концевой отдел
неразветвлен)
МЕХАНИЗМЫ ВЫДЕЛЕНИЯ СЕКРЕТА
МЕРОКРИНОВЫЙ
АПОКРИНОВЫЙ (МИКРО-, МАКРО)
ГОЛОКРИНОВЫЙ
46.
47.
Лекция окончена.Благодарю за
внимание.