








































 суммация в возбуждающих синапсах")













 biology
biologySimilar presentations:










Общая физиология ЦНС. Рефлекторный принцип деятельности. Торможение
1. Общая физиология ЦНС Рефлекторный принцип деятельности. Торможение.
Профессор Берг М.Д.2.
План лекции1. Структура центральной нервной системы, нейроглия и её функции.
2. Виды и функции нейронов, функциональная характеристика
отростков. Механизм возбуждения нейрона.
3. Взаимоотношения нейронов с другими клетками. Виды и структура
синапсов.
4. Механизм работы возбуждающих и тормозящих синапсов.
5. Центральные медиаторы и хеморецепторы (ионотропные и
метаботропные).
6. Рефлекс, рефлекторная дуга. Характеристика звеньев рефлекторной
дуги. Моно и полисинаптические рефлексы.
7. Нервный центр, межнейрональные связи, функции центров.
Особенности передачи информации в нервных центрах.
8. Характеристика процесса возбуждения.
9. Характеристика процесса торможения, классификация видов,
значение.
10.Координация рефлекторных процессов. Механизмы, связанные с
торможением, механизмы обратной связи, доминанты,
субординации.
Профессор Берг М.Д.
3.
Центральная нервная система представленаголовным и спинным мозгом. Основные клетки –
нейроны разных видов. Общее количество у
человека - 13,6 млрд.
Кроме того, - глиальные клетки и межклеточное вещество,
составляющее около 20% объема мозга.
Глиальные клетки трех видов: олигодендроциты, астроциты,
микроглиальные клетки. Функции глии многообразны:
опорная, трофическая, иммунная, миелинообразуюшая,
дренажная, участие в синаптической пластичности и
формировании долговременной памяти.
Олигодендроциты своими отростками образуют: а)
миелиновую оболочку аксонов, толщину которой они
контролируют с учетом функциональной активности нейрона,
б) глимфатическую систему для управления притоком и
оттоком воды и веществ в мозг и из мозга.
Профессор Берг М.Д.
4.
У астроцитов, кроме трофической и энергетической функций,выявлена важная роль в управлении толщиной миелиновой
оболочки и шириной перехватов Ранвье в результате
блокирования тромбина, расщепляющего белки, связывающие
между собой листки миелина в области перехватов Ранвье.
Астроциты, лежащие вблизи синапсов, влияют на синаптическую
передачу, поглощая или выделяя нейромедиатор. Это пути
участия астроцитов глии в формировании долговременной
памяти.
Микроглиальные клетки выполняют иммунную функцию –
фагоцитируют ненужные молекулы, потенциально вредные
вещества, обломки погибших клеток. Никогда не покидают
пределов мозга. Очень важная функция микроцитов – участие в
синаптической пластичности: уничтожают лишние синапсы,
помогают создавать новые синапсы, прокладывают для них путь
в межклеточном пространстве, «проедая» туннель, влияют на
количество дендритных шипиков, т.е. редактируют нейронные
сети; контролируют также качество миелиновой оболочки.
Профессор Берг М.Д.
5.
Красные точки – клетки микроглииспинного мозга мыши.
Профессор Берг М.Д.
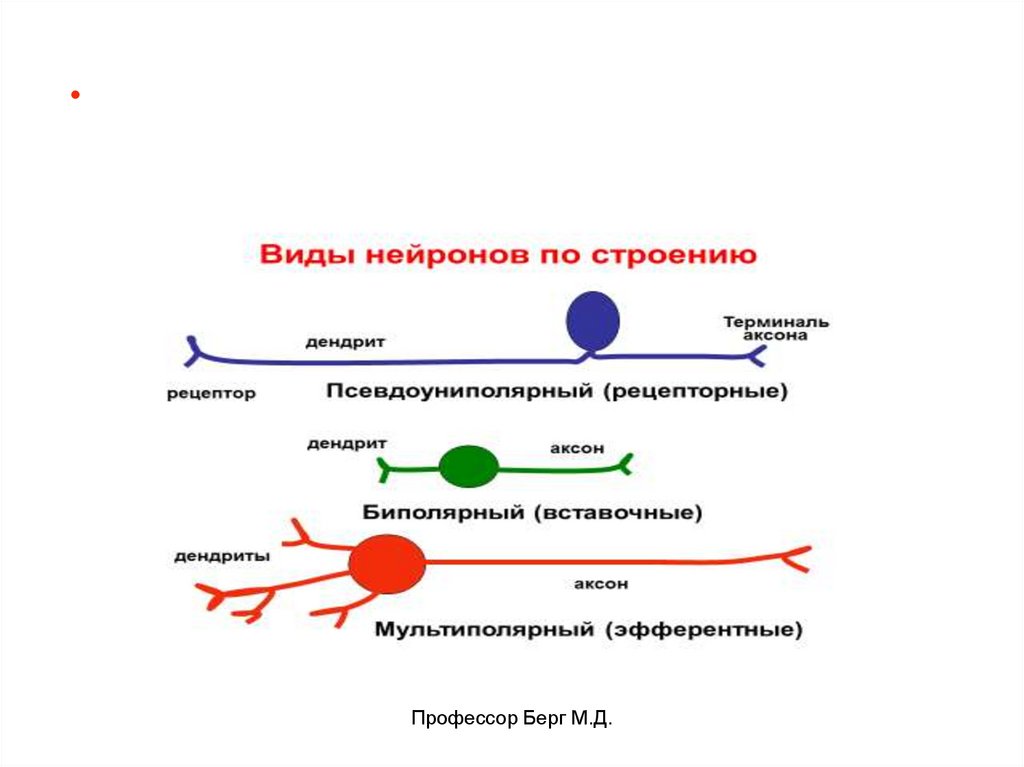
6. Виды и функции нейронов, функциональная характеристика отростков.
Профессор Берг М.Д.7.
Нейрон – структурно-функциональная единицанервной системы.
Функции нейронов: восприятие, кодирование,
передача, переработка информации, декодирование
в психонервный процесс, запоминание.
Нейрон имеет тело и 2 вида отростков:
• аксон (всегда один),
• дендриты.
Дендриты проводят возбуждение к телу нейрона,
аксон – от тела нейрона к другим клеткам.
Возбуждение в нервных волокнах возникает и
проводится благодаря потенциалзависимым Na+
каналам плазматической мембраны – Nav1.3.
(тетродотоксин, рыба фугу).
В безмиелиновых волокнах каналы располагаются
по всей мембране с плотностью 110-120 на 1 мкм2, в
миелиновых – в зоне
перехватов Ранвье.
Профессор Берг М.Д.
8.
Скорость проведения по безмиелиновым волокнам –0,5 - 3 м/с, миелиновым – 3 - 120 м/с.
По скорости проведения ПД нервные волокна делят
на 3 группы:
• А, V= 20-120 м/с
• В, V= 3-20 м/с
• С, V= 0,5-3 м/с
Схема проведения возбуждения по
нервным волокнам
ПД
ПД
Профессор Берг М.Д.
9.
Виды нейронов:• афферентные (рецепторные, чувствительные),
воспринимают, кодируют и передают информацию от
рецепторов к вставочным или двигательным
нейронам.
• вставочные (промежуточные, интернейроны)
связывают между собой афферентные и
эфферентные нейроны.
• эфферентные (исполнительные) нейроны 2-х видов:
- двигательные (моторные, соматические),
- вегетативные (преганглионарные и постганглионарные нейроны симпатической и парасимпатической
систем).
• ассоциативные нейроны (обеспечивают связь
между нервными центрами).
Профессор Берг М.Д.
10.
Законы проведения:1. ЗАКОН БЕЗДЕКРЕМЕНТНОГО ПРОВЕДЕНИЯ. ПД не меняет
свою амплитуду на протяжении всего пути.
безмиелиновое
Профессор Берг М.Д.
миелиновое
11.
ЗАКОН ИЗОЛИРОВАННОГО ПРОВЕДЕНИЯ2. В нервном пучке ПД не передается с одного
волокна на соседнее.
Профессор Берг М.Д.
12.
Профессор Берг М.Д.
13. ЭФФЕРЕНТНЫЕ НЕЙРОНЫ
двигательный нейрон переднихрогов спинного мозга
АХ
АХ
ВЕГЕТАТИВНЫЕ
НЕЙРОНЫ
Профессор Берг М.Д.
АХ
или
НА
С
К.
М
Ы
Ш
Ц
А
Г
Л.
М
Ы
Ш
Ц
А
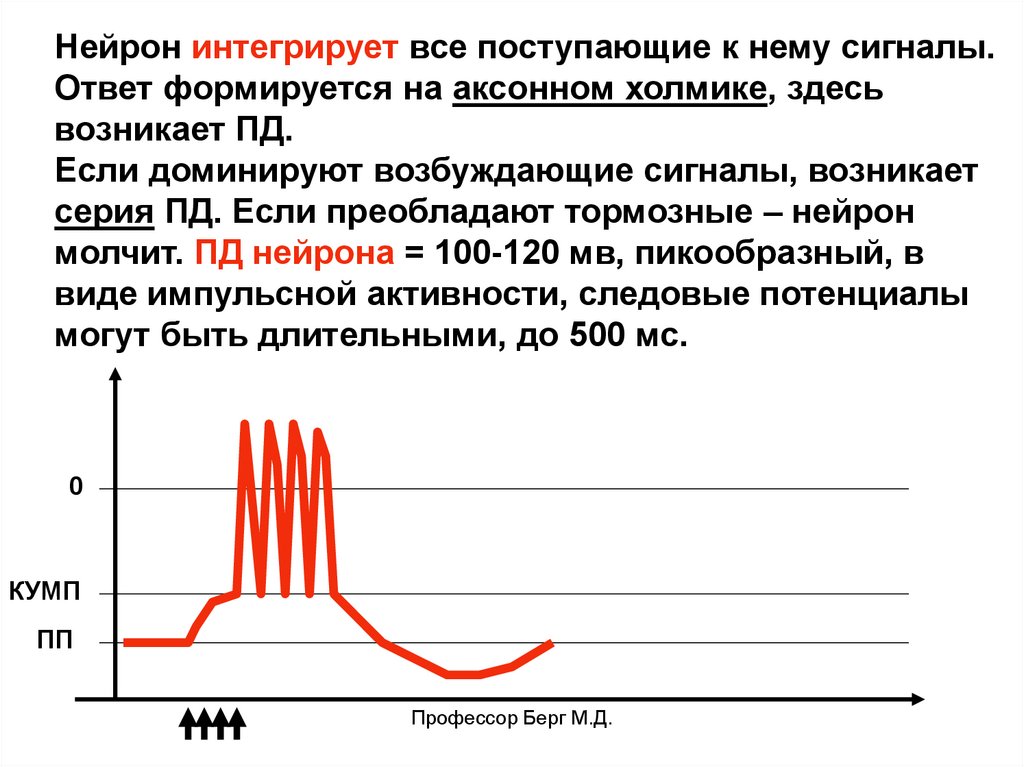
14.
Нейрон интегрирует все поступающие к нему сигналы.Ответ формируется на аксонном холмике, здесь
возникает ПД.
Если доминируют возбуждающие сигналы, возникает
серия ПД. Если преобладают тормозные – нейрон
молчит. ПД нейрона = 100-120 мв, пикообразный, в
виде импульсной активности, следовые потенциалы
могут быть длительными, до 500 мс.
0
КУМП
ПП
Профессор Берг М.Д.
15.
Виды межнейрональных структур:1. Нейронные цепи:
• Рефлекторные дуги,
• Проводящие пути.
2. Нейронные сети:
• локальные,
• иерархические (расположены на разных уровнях
ЦНС).
Профессор Берг М.Д.
16.
Взаимоотношения нейронов сдругими клетками.
Виды и структура синапсов.
Контакты аксона нейрона с другими клетками
называют синапсом. Они могут быть: 1) с другими
нейронами (центральные синапсы и синапсы
вегетативных ганглиев) или
2) с рабочими клетками – периферические синапсы:
• нервно-мышечные синапсы со скелетными
мышцами,
• вегетативные синапсы с гладкими мышцами и
железистыми клетками.
Профессор Берг М.Д.
17.
По механизму работы синапсы могут бытьвозбуждающими и тормозящими.
В структуре синапса выделяют:
1. Пресинаптическую часть. Она представлена
утолщением терминали.
2. Синаптическую щель.
3. Постсинаптическую часть – часть мембраны
клетки, с которой образован синапс.
Синаптическая щель в химическом синапсе – 20-50
нм. Здесь есть ферменты для расщепления
медиатора – ферменты-дворники.
Профессор Берг М.Д.
18.
Структура синапсатерминаль аксона
медиатор
Са++
Пресинаптическая
мембрана
синаптическая щель
хеморецептор
Постсинаптическая
мембрана
Внесинаптическая мембрана
Мышечное волокно
Профессор Берг М.Д.
19.
Механизм работы синапса(на примере нервно-мышечного синапса)
ПД
терминаль аксона
мотонейрона
медиатор АХ
Са++
Пресинаптическая
мембрана
синаптическая щель
ПД
Н-холинорецептор
Постсинаптическая
мембрана
ЛО
Мышечное волокно
Профессор Берг М.Д.
20.
Механизм работы возбуждающих синапсов:1. ПД распространяется по терминали и открывает п/з
Са++ -каналы пресинаптической мембраны.
2. Са++ входит в терминаль, взаимодействует с
мембраной синаптических пузырьков, которые
сливаются с пресинаптической мембраной, образуя
канал.
3. Медиатор изливается в синаптическую щель
(экзоцитоз), диффундирует к постсинаптической
мембране.
4. Медиатор взаимодействует с хеморецепторами
постсинаптической мембраны, это приводит к
открытию х/з ионных каналов для Na+, он входит в
клетку, возникает ЛО или возбуждающий постсинаптический потенциал (ВПСП).
Профессор Берг М.Д.
21.
5. Если ЛО достигает КУМП, открываются п/з Na+каналы внесинаптической мембраны, возникает ПД (в
нейроне – это мембрана аксонного холмика).
6. Медиатор инактивируется ферментами, которые
очищают рецепторы постсинаптической мембраны,
рецепторы готовы к новому взаимодействию с
медиатором.
Ферменты-дворники специализированы к работе с
определенным медиатором (ацетилхолин + холестеринэстераза
= холин + уксусная к-та; норадреналин + моноаминоксидаза или катехол-Ометилтрансфераза = Н2О2 + NН3 + альдегиды).
Продукты метаболизма вновь используются для
ресинтеза медиатора или удаляются.
Не использованный медиатор может подвергаться
обратному захвату. Харуки Мураками «Подземка». Аум Сенрике – зарин, уменьшение
эффектов холинэстеразы.
На синаптическую передачу расходуется 0,5-1,0 мс –
синаптическая задержка.
Профессор Берг М.Д.
22.
Механизм работы тормозящих синапсов.Первые этапы (экзоцитоз):
1. 2. 3. как и в возбуждающем синапсе.
4. Медиатор взаимодействует с хеморецепторами
постсинаптической мембраны, это приводит к
открытию ионных каналов для К+ или Cl-. К+ выходит
из клетки через постсинаптическую мембрану (Clвходит в клетку).
5. Это вызывает гиперполяризацию постсинаптической мембраны. Возбудимость клетки снижается и она
не реагирует на возбуждающие сигналы, т.е.
тормозится.
Более детально этапы жизни везикул с медиатором и
механизм экзоцитоза представлен на рисунках ниже.
Профессор Берг М.Д.
23.
Этапы жизни везикулОтпочковывание
пузырьков от
эндосомы
Заполнение
синаптических
пузырьков
медиатором
Транспорт
пузырьков
к местам
освобожде
ния
Подход к
мембране
Подготовка к
слиянию
Слияние и опорожнение
пузырька (экзоцитоз)
Профессор Берг М.Д.
24.
Этапы экзоцитозаПрофессор Берг М.Д.
25.
Виды центральных синапсов.Аксон может контактировать с
• дендритом (аксо-дендритный синапс),
• телом (аксо-соматический синапс)
• аксоном другой клетки (аксо-аксональный синапс).
Профессор Берг М.Д.
26.
Аксо-дендритные синапсы всегда возбуждающие,аксо-соматические – возбуждающие и тормозящие,
аксо-аксональные – тормозящие.
На одном нейроне – до 6000 синапсов.
Строение синапса типичное: окончание терминали,
синаптическая щель, постсинаптическая часть.
Профессор Берг М.Д.
27.
Центральные медиаторы ихеморецепторы
Каждый нейрон синтезирует свойственный ему
медиатор.
Виды медиаторов:
ацетилхолин (АХ), норадреналин (НА), адреналин (А),
серотонин, гистамин, вещество «Р»,
эндорфины, энкефалины,
аденозин, АТФ и др. возбуждающие медиаторы,
глицин, гаммааминомасляная кислота (ГАМК) –
тормозные медиаторы.
Профессор Берг М.Д.
28.
Виды хеморецепторов.Каждому медиатору на постсинаптической мембране
соответствует свой хеморецептор.
Его называют по виду медиатора:
АХ – холинорецептор, норадреналин – адренорецептор и т.д.
По механизму работы:
1. Ионотропные хеморецепторы.
Они являются ионными каналами. При взаимодействии с медиатором белок-рецептор изменяет
форму («открывается» канал) и пропускает ион.
2. Метаботропные хеморецепторы.
Медиатор взаимодействует с белком-рецептором и
запускает цепочку событий, приводящих к открытию
ионных каналов с помощью вторичных посредников.
Эти рецепторы работают с участием G-белка
(Нобелевская премия Профессор
2014 года).
Берг М.Д.
29.
медиаторИонотропные хеморецепторы
Профессор Берг М.Д.
30.
Метаботропные хеморецепторыМедиатор (АХ)
М-холинорецептор
G
Ca++
ДГ
Фосфолипаза С
Запускает биохимические
и другие реакции клетки
Профессор Берг М.Д.
31. Рефлекторный принцип деятельности ЦНС
Профессор Берг М.Д.32.
Рефлекс - ответная реакция организма на раздражениес участием ЦНС.
Сигнал может поступить из внешней или внутренней
среды. Рефлекс осуществляется по рефлекторной
дуге. Это – цепь нейронов: афферентный, вставочный,
эфферентный. В ней выделяют звенья:
1) рецептор. Виды:
• Экстерорецепторы воспринимают сигналы из
внешней среды (свет, звуки, запахи, прикосновение,
температура и др.)
• Интерорецепторы воспринимают сигналы из внутренней среды организма (давление крови, рН, рО2, рСО2, и
др.).
• Проприорецепторы реагируют на сокращение и
расслабление мышц, движение в суставах.
Скопление рецепторов – рецептивное поле, имеет
размеры от 0,02 до 20 Профессор
см. Берг М.Д.
33.
Чувствительные нейроны.Аксоноподобный дендрит чувствительных
нейронов заканчивается рецептором, который
реагирует на раздражение.
Профессор Берг М.Д.
34.
По строению и механизму возбуждения рецепторыклассифицируются на:
• первичночувствующие,
• вторичночувствующие.
Профессор Берг М.Д.
35.
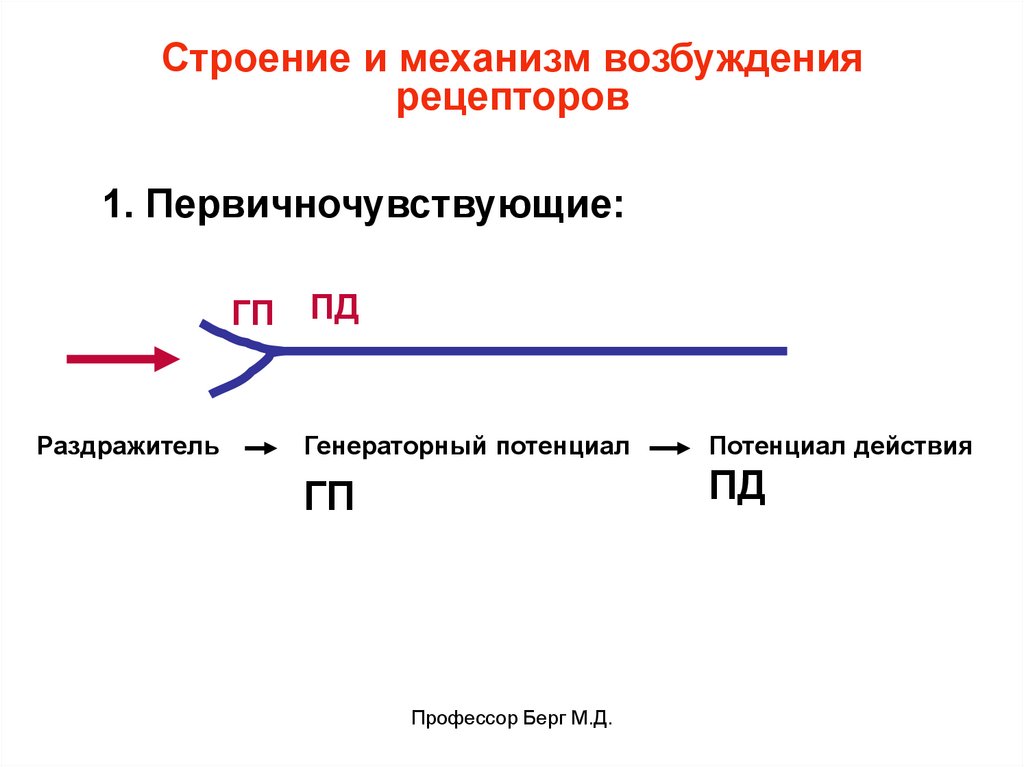
Строение и механизм возбуждениярецепторов
1. Первичночувствующие:
ГП
Раздражитель
ПД
Генераторный потенциал
Потенциал действия
ГП
ПД
Профессор Берг М.Д.
36. 2.Вторичночувствующие
Секреция медиатораРП
Специализированная
волосковая клетка
ГП
ПД
Нервное окончание чувствительного
нейрона
Стимул --- РП (рецепторный потенциал) --секреция медиатора --- ГП --- ПД
Профессор Берг М.Д.
37.
Дальнейшие звенья рефлекторной дуги:2) Афферентный путь. От рецептора до ЦНС. V=0,5-70
м/с.
3) Нервный центр (спинальный, бульбарный и др. – по
отделам мозга).
4) Эфферентный путь двигательный (соматический)
или вегетативный.
5) Рабочий орган (мышца, железа).
При функциональном или анатомическом выключении
любого звена рефлекс не возникает.
Время, необходимое для проведения возбуждения по
рефлекторной дуге, называют временем рефлекса
(0,1-0,3 с).
Профессор Берг М.Д.
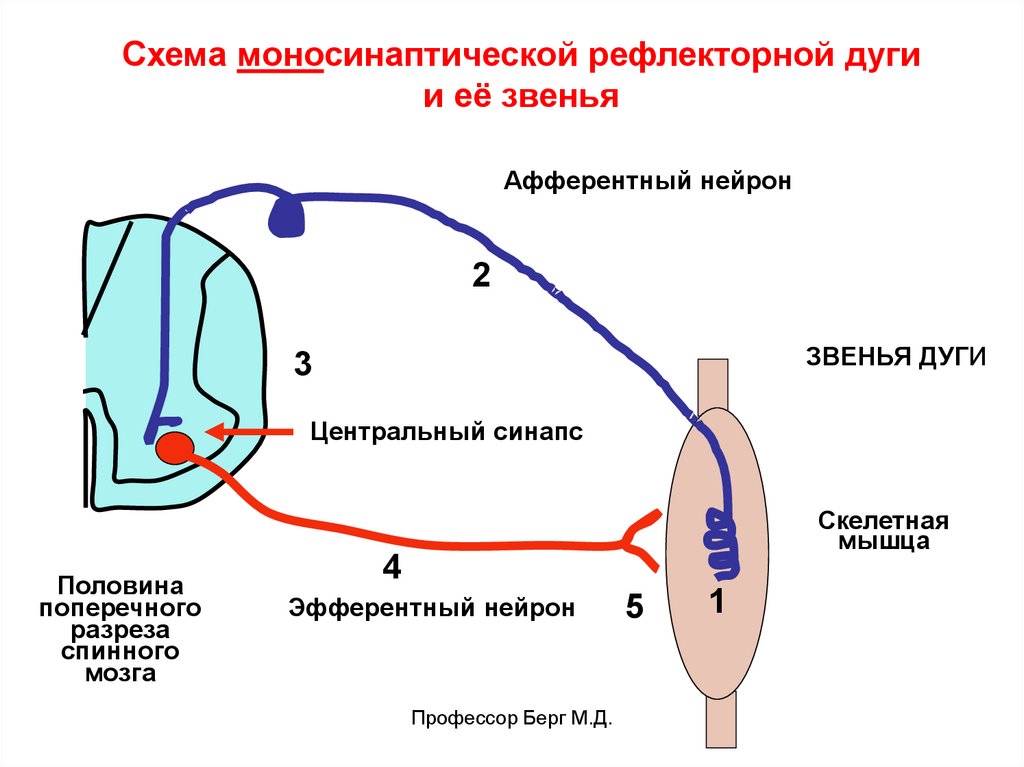
38.
Схема моносинаптической рефлекторной дугии её звенья
Афферентный нейрон
2
ЗВЕНЬЯ ДУГИ
3
Центральный синапс
Половина
поперечного
разреза
спинного
мозга
Скелетная
мышца
4
Эфферентный нейрон
Профессор Берг М.Д.
5
1
39.
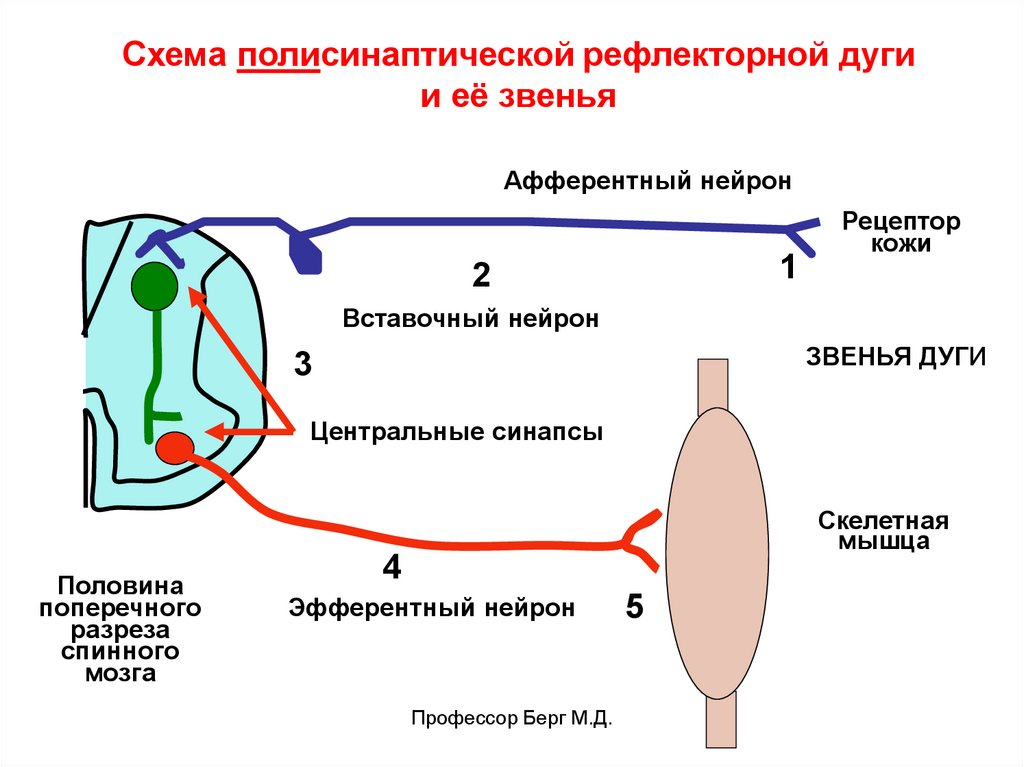
Схема полисинаптической рефлекторной дугии её звенья
Афферентный нейрон
1
2
Рецептор
кожи
Вставочный нейрон
ЗВЕНЬЯ ДУГИ
3
Центральные синапсы
Половина
поперечного
разреза
спинного
мозга
Скелетная
мышца
4
Эфферентный нейрон
Профессор Берг М.Д.
5
40. НЕРВНЫЙ ЦЕНТР. ОСОБЕННОСТИ ПЕРЕДАЧИ ИНФОРМАЦИИ В НЕРВНЫХ ЦЕНТРАХ
Нервный центр – группа нейронов, отвечающая затот или иной рефлекс.
Центр может быть образован локальными или
иерархическими сетями нейронов.
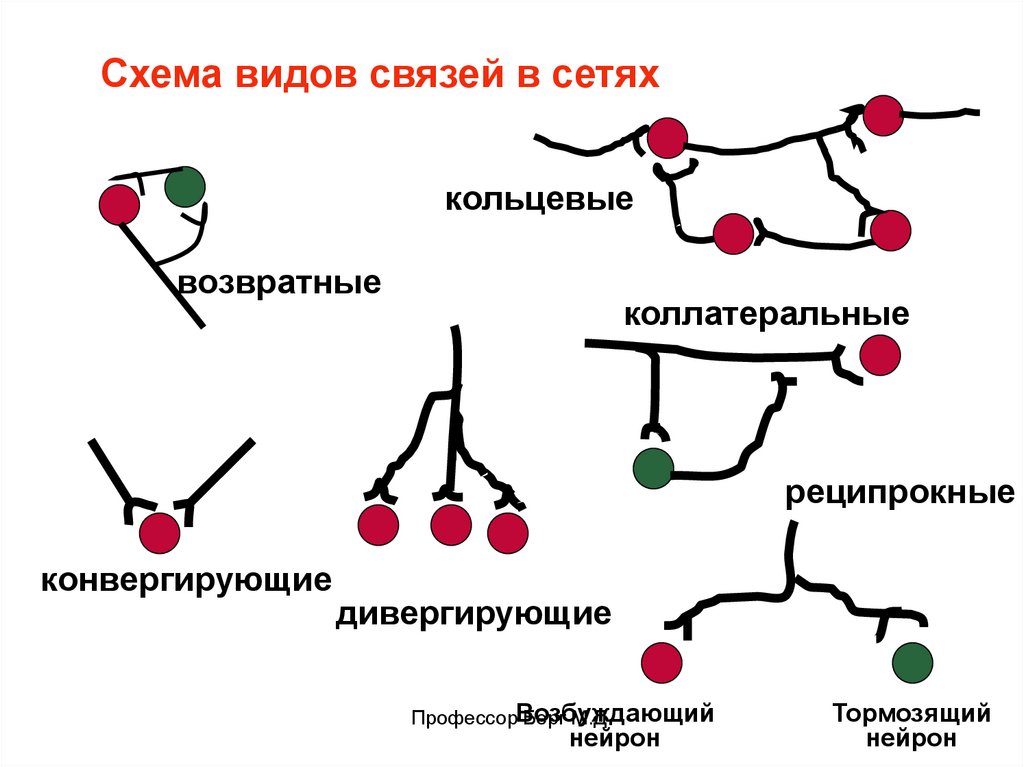
Виды связей в сетях: конвергирующие, дивергирующие, коллатеральные, возвратные, реципрокные,
кольцевые.
Профессор Берг М.Д.
41.
Схема видов связей в сетяхкольцевые
возвратные
коллатеральные
реципрокные
конвергирующие
дивергирующие
ПрофессорВозбуждающий
Берг М.Д.
нейрон
Тормозящий
нейрон
42.
ФУНКЦИИ СЕТЕЙ (НЕРВНЫХ ЦЕНТРОВ):• Регуляция входа информации (фильтрация –
очищение сигнала, усиление или ослабление,
контрастирование) и её анализ.
• Выбор конечного пути рефлекса, запуск и
реализация врожденных или приобретенных
программ, их модулирование.
• Обучение и сохранение (запоминание) информации.
Профессор Берг М.Д.
43.
СВОЙСТВА ЦЕНТРАЛЬНЫХСИНАПСОВ и НЕРВНЫХ ЦЕНТРОВ:
1. Одностороннее проведение сигнала.
2. Задержка проведения (один синапс – 0,3 мс-1мс).
3. Суммация возбуждения или торможения.
4. Трансформация ритма (изменение ритма: на выходе
из центра ритм больше или меньше, чем на входе).
5. Низкая лабильность (менее 100 Гц).
6. Повышенная утомляемость.
6. Чувствительность к недостатку кислорода и
нейротропным ядам (без кислорода могут жить:
КБП – 5, ствол – 15, спинной мозг – 30 мин.).
7. Посттетаническая потенциация
(облегчение проведения сигнала после предварительной работы центра за счет увеличения Са++, медиатора,
ВПСП; проявляется в уменьшении времени рефлекса,
Профессор Берг М.Д.
лежит в основе обучения).
44.
8. Синаптическая депрессия (ухудшениесинаптической передачи в центре после
предварительной работы за счет уменьшения
чувствительности постсинаптической мембраны изза снижения активности ферментов-дворников или
других причин; увеличение времени рефлекса).
9. Последействие (ответ продолжается при
отсутствии раздражения за счет реверберации
нервных импульсов по замкнутым цепям).
10. Тонус нервных центров (активность в покое за
счет нервной или гуморальной стимуляции).
11. Пластичность нервных центров (способность
приспосабливаться при изменении условий или при
утрате части нейронов).
Профессор Берг М.Д.
45.
В нервных центрах работают 2процесса: возбуждение и торможение.
Возбуждение – это процесс, возникающий в ответ на
раздражение. Характеризуется появлением или
усилением ответной реакции организма.
Связан с деятельностью возбуждающих нейронов,
которые синтезируют и выделяют в синапсах
возбуждающие медиаторы. ПД возникает в результате
суммирования ВПСП.
Виды суммирования:
- пространственная суммация
(суммируются ВПСП разных синапсов одного
нейрона),
- последовательная суммация (суммируются ВПСП,
возникающие последовательно одном и том же
синапсе).
Профессор Берг М.Д.
46.
При суммации ВПСП достигает критическогоуровня мембранного потенциала (КУМП) аксонного
холмика и нейрон генерирует серию ПД.
Возбуждающие медиаторы центральных синапсов:
АХ, НА, А, глютамат, серотонин, ДОФА, вещество Р
и др.
Профессор Берг М.Д.
47. Пространственная суммация в возбуждающих синапсах
ПДПД
ПД
нейрон
ПДн
0
Екр
Епп
Профессор Берг М.Д.
48. Последовательная (временная) суммация в возбуждающих синапсах
ПДнейрон
ПДн
0
Екр
Епп
Профессор Берг М.Д.
49.
Характеристика процесса трможенияТорможение – второй процесс, который работает в
ЦНС. Он тоже возникает в ответ на раздражение.
Проявляется отсутствием или снижением ответной
реакции на раздражение.
Связан с работой тормозных нейронов, которые
синтезируют и выделяют в окончаниях аксонов
тормозные медиаторы.
Их два: глицин и ГАМК.
Профессор Берг М.Д.
50.
Классификация центрального торможения:1.Пресинаптическое
2.Постсинаптическое:
• Прямое
• Возвратное
• Реципрокное
• Латеральное
Постсинаптическое торможение
снижает возбудимость постсинаптической
мембраны.
Профессор Берг М.Д.
51.
Пресинаптическое торможение связано с работой аксоаксональных синапсов (между аксоном тормозногонейрона и терминалью аксона возбуждающего нейрона).
Итог – снижение секреции возбуждающего медиатора.
Механизм:
Тормозной медиатор --- диффузия ионов К+ или СI- --гиперполяризация пресинаптической мембраны --снижение чувствительности (блокирование) Са++ каналов.
Профессор Берг М.Д.
52.
СХЕМА ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯСа++
ПД
Са
ПД
Са++
глицин
ПД
Профессор Берг М.Д.
ВПСП
53.
Пример рефлекторной дуги с пресинаптическимторможением
Профессор Берг М.Д.
54.
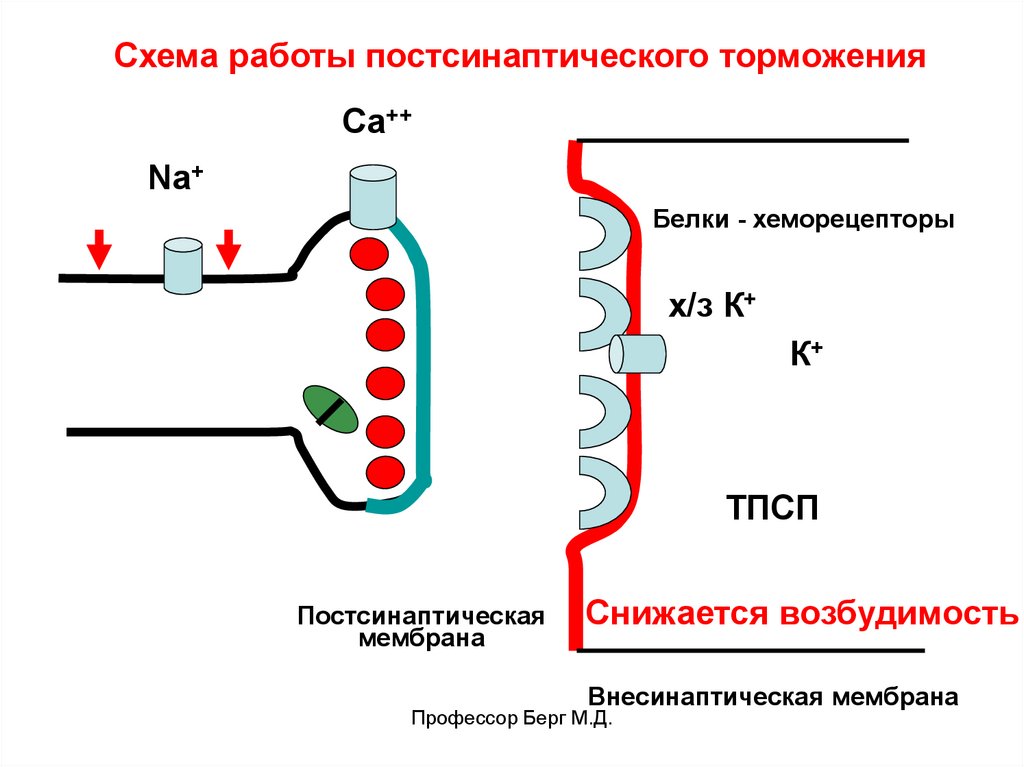
Схема работы постсинаптического торможенияСа++
Na+
Белки - хеморецепторы
х/з К+
К+
ТПСП
Постсинаптическая
мембрана
Снижается возбудимость
Внесинаптическая мембрана
Профессор Берг М.Д.
55.
Виды постсинаптического торможения:1) Прямое – возбуждение сразу поступает на
тормозной нейрон.
2) Реципрокное – возбуждение поступает
одновременно на возбуждающий и тормозящий
нейроны.
3) Возвратное – возбуждение поступает по
коллатерали мотонейрона на тормозной нейрон
Реншоу и снижает возбудимость этого же
мотонейрона.
4) Латеральное – возбуждение поступает по
коллатералям на тормозные нейроны и блокирует
проведение по параллельным путям.
Профессор Берг М.Д.
56.
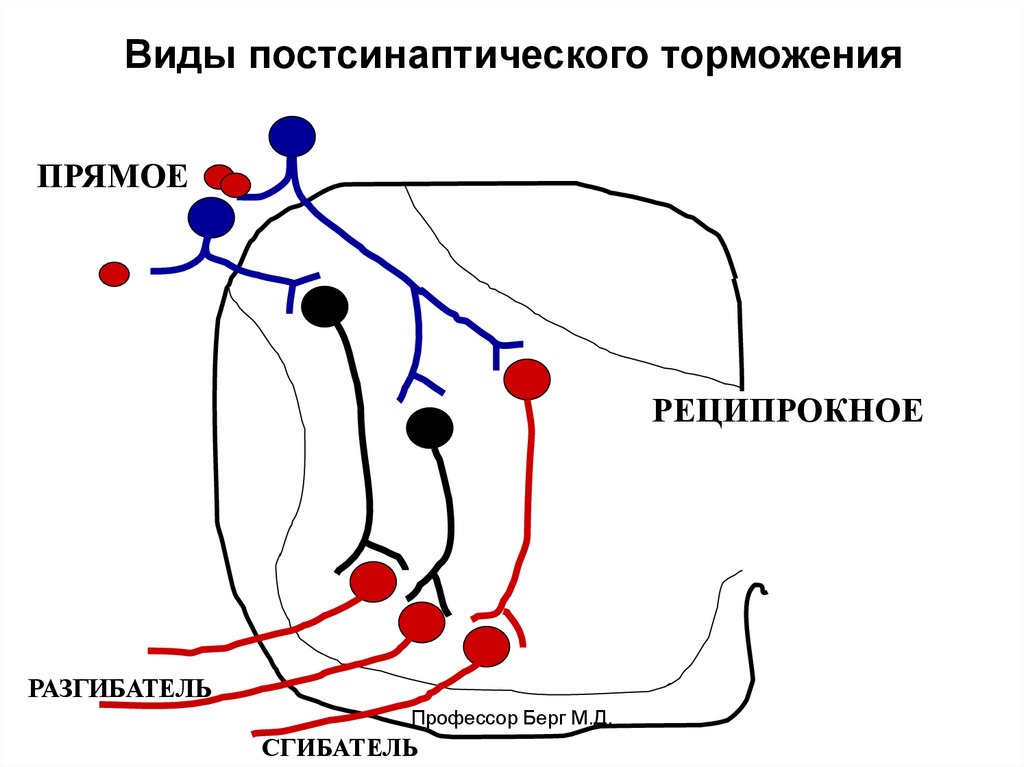
Виды постсинаптического торможенияПРЯМОЕ
РЕЦИПРОКНОЕ
РАЗГИБАТЕЛЬ
Профессор Берг М.Д.
СГИБАТЕЛЬ
57.
Виды постсинаптического торможенияВОЗВРАТНОЕ
ЛАТЕРАЛЬНОЕ
Клетка Реншоу
Профессор Берг М.Д.
58.
Механизмы координациирефлекторной деятельности:
Центральное торможение нужно для координации
рефлекторной деятельности наряду со специальными механизмами координации.
Профессор Берг М.Д.
59.
Механизмы координациирефлекторной деятельности:
1. Взаимодействие возбуждения и торможения на
одном нейроне путем суммации ВПСП и ТПСП.
2. Взаимодействие возбуждения и торможения на
уровне нервных центров за счет реципрокного
торможения.
Профессор Берг М.Д.
60.
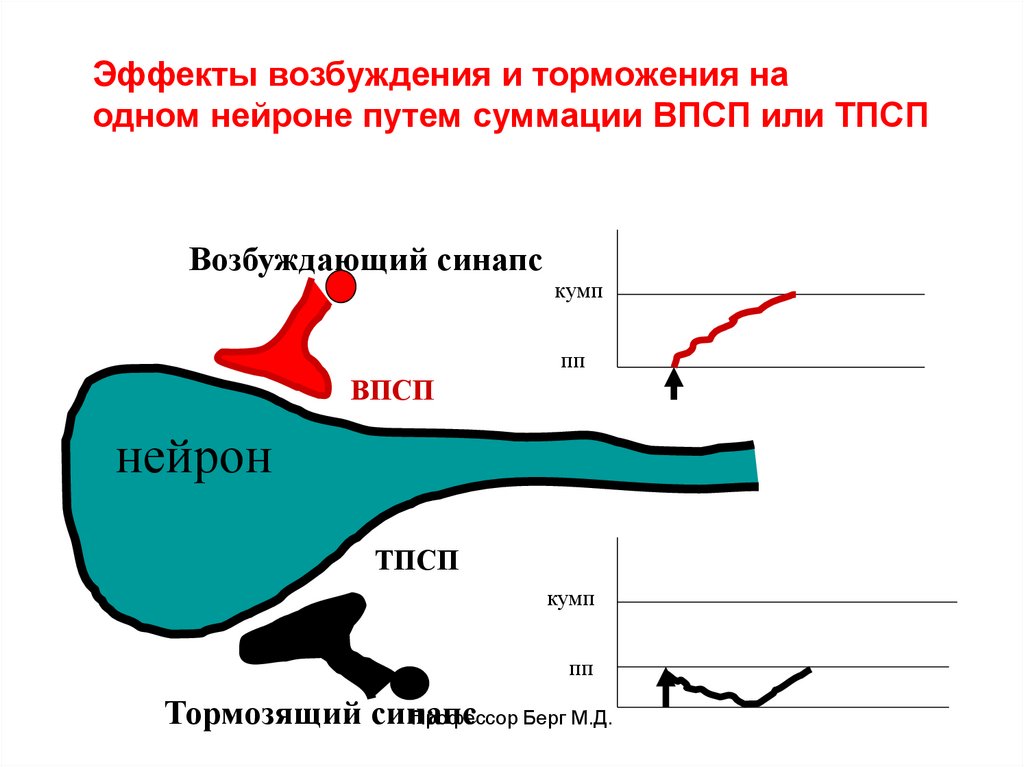
Эффекты возбуждения и торможения наодном нейроне путем суммации ВПСП или ТПСП
Возбуждающий синапс
кумп
пп
ВПСП
нейрон
ТПСП
кумп
пп
Тормозящий синапс
Профессор Берг М.Д.
61.
Взаимодействие возбуждения и торможения наодном нейроне путем суммации ВПСП и ТПСП
Возбуждающий синапс
кумп
пп
ВПСП
нейрон
ТПСП
Тормозящий синапс
Профессор Берг М.Д.
62.
Взаимодействие возбуждения и торможения науровне нервных центров обеспечивает связи между
нейронными сетями, которые управляют
функциями правой и левой частей тела, верхними и
нижними конечностями, мышцами сгибателями и
разгибателями одной стороны. Эти связи
реципрокные, т.е. антагонистические, работают
через тормозные нейроны постсинаптического
торможения.
Профессор Берг М.Д.
63.
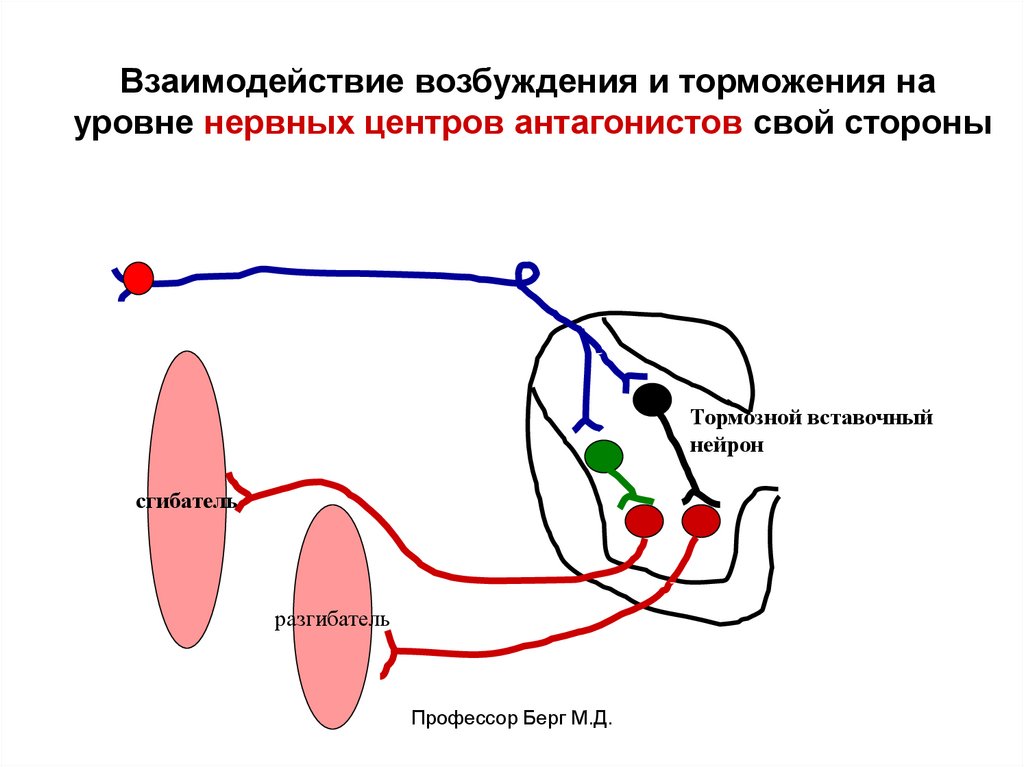
Взаимодействие возбуждения и торможения науровне нервных центров антагонистов свой стороны
Тормозной вставочный
нейрон
сгибатель
разгибатель
Профессор Берг М.Д.
64.
3. Использование механизма обратной связи(положительной или отрицательной).
Отрицательная – уменьшает рефлекторный ответ,
положительная – усиливает.
Механизм обратной связи разработан П.К.Анохиным.
Обратная связь – это связь рабочего органа с
нервными центрами.
За счет нее:
• нервные центры информируются о выполнении или
невыполнении задания,
• изменяется уровень активности нервных центров.
Выделяют положительную и отрицательную
обратную связь (обратная афферентация
усиливающая или ослабляющая функции центра).
Профессор Берг М.Д.
65.
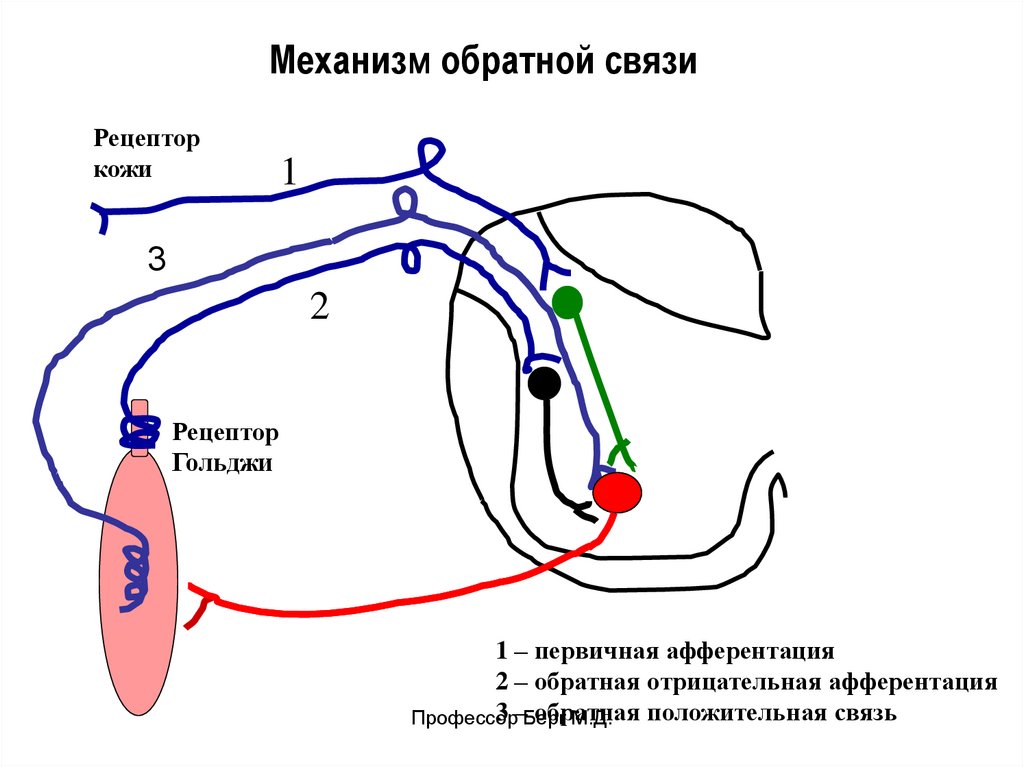
Механизм обратной связиРецептор
кожи
1
3
2
Рецептор
Гольджи
1 – первичная афферентация
2 – обратная отрицательная афферентация
3 –Берг
обратная
положительная связь
Профессор
М.Д.
66.
4. Принцип доминанты (А.А.Ухтомский) –формирование господствующего нервного центра,
подчиняющего себе работу других центров для
удовлетворения текущей потребности.
Свойства:
- повышенная возбудимость,
- стойкость возбуждения,
- способность притягивать и суммировать
возбуждения, идущие к другим центрам,
- инерция возбуждения.
5. Принцип субординации
(подчинение нижележащих центров вышележащим).
Например, центры мочеиспускания и дефекации
подчиняются корковым центрам, формируется
подчинение с 2-3-летнего возраста.
Профессор Берг М.Д.
67.
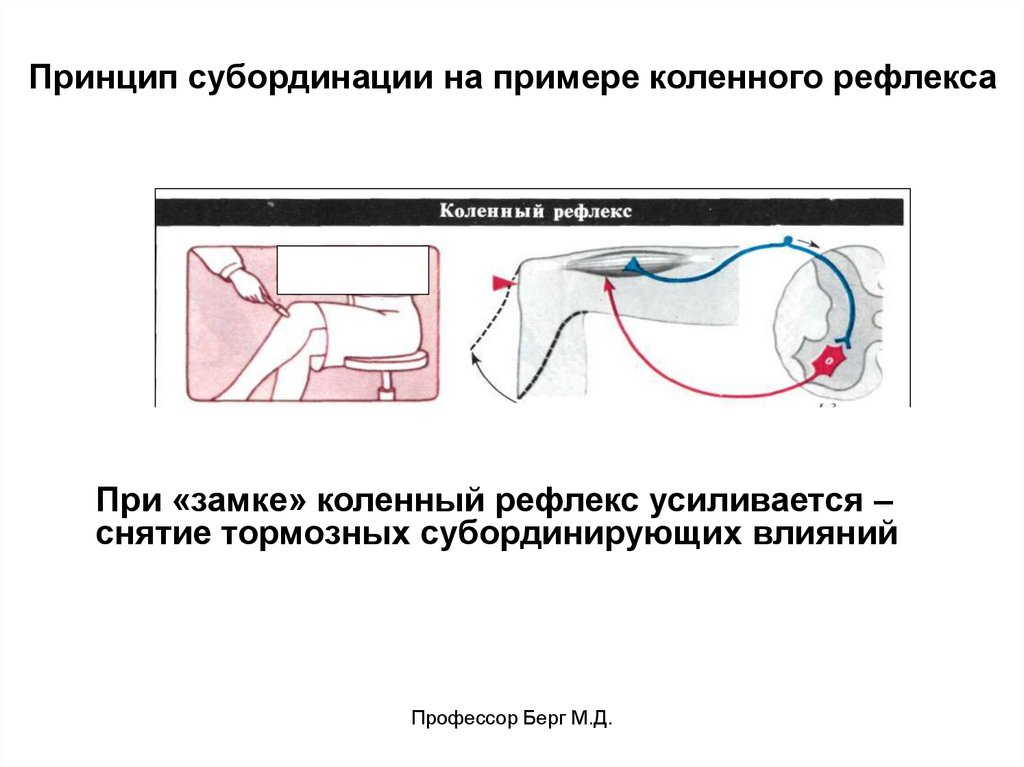
Принцип субординации на примере коленного рефлексаПри «замке» коленный рефлекс усиливается –
снятие тормозных субординирующих влияний
Профессор Берг М.Д.