















 2. экстирпация (удаление) 3. трансплантация 4. фистульный 5.")










 Перенос ионов натрия и калия через мембрану. Принцип работы натрий-")
. Натрий- кальциевый обменник")
















 каналов")

 управляемых ионных каналов")







")



 итоговая движущая сила (V- Е)")


 biology
biologySimilar presentations:
")









Физиология возбудимых тканей. Ионные каналы, строение, классификация, способ активации, прикладные аспекты
1. Лекция 1
Физиология возбудимых тканейИонные каналы, строение, классификация,
способ активации, прикладные аспекты
2. Кафедра нормальной физиологии КГМУ
Основана в 1805 годуhttp://kgmu.kcn.ru/physiology
3. Курс нормальной физиологии
ЛекторНигматуллина Разина
Рамазановна
доктор биологических наук,
профессор
4. История кафедры
5. И.П.Павлов, 1927 год
“…оценивая труды Казанскихфизиологов c полным основанием
можно говорить о целой
физиологической школе…”
И.П.Павлов, 1927 год
6.
Ф. В. Овсянников“Состояние учебных кабинетов и
лабораторий вообще превзошло
мое ожидание...
Физиологический институт, …
оказались в избытке снабженными
инструментами, аппаратами и
другими вспомогательными
средствами для научного
преподавания и для упражнения
учащихся…
Университет может существовать и
постоянно преуспевать своими
собственными средствами, потому
что в нем нет недостатка в дельных
и усердных преподавателях.”
К. Бэр
7.
“Число работ, вышедших излаборатории профессора
Миславского чрезвычайно
велико.
В этих работах затронуты большие
существенные вопросы
современной физиологии, и они
в своей совокупности
составляют настоящую
физиологическую библиотеку…”
“В заграничных лабораториях
Миславский не только учился у своих
товарищей и друзей, но и являлся
учителем, демонстрируя опыты и
особенность их постановки в его
Казанской лаборатории.”
Миславский Н.И.
Самойлов А.Ф.
8. Павлов И.П.
“Хорошая привычка физиологическидумать явится драгоценным
пособием к чисто медицинскому
знанию...
Без этой привычки врач будет прямо
жалок около больного.
Я убежден, что только благодаря
частому обмену мнений между
физиологом и врачом цель
физиологической науки и
медицинского искусства будет
быстрейшим и важнейшим образом
достигнута.”
Павлов И.П.
9.
А.Ф. Самойлов10. Академик АМН СССР В.В. Парин
” Сюда, как паломники вМекку съезжались со
всей России физиологи,
чтобы научиться
работать с
Эйнтховенским струнным
гальванометром - святая
святых лаборатории.
То был один из первых
электрокардиографических
аппаратов, казавшийся
тогда чудом техники.”
Академик АМН СССР В.В. Парин
11. Высказывание Самойлова
- … во всех случаях, когда возбуждение переходит отодной клетки на другую, будь то в синапсе или на
границе между нервным и мышечным волокном, такие
особенности перехода, как одностороннее
проведение, временная задержка, можно объяснить,
если принять, что из двух соприкасающихся клеток
одна приобрела способность вырабатывать
раздражающее вещество, а другая – способность
реагировать на него
12.
“Выводы:1. В симпатической системе
переход возбуждения с одного
нейрона на другой совершается при
участии химических агентов.
2. Участие химического фактора
обязательно при переходе
возбуждения с одной клетки на
другую и в других частях нервной
системы.
Кибяков А.В.
3. Функциональные свойства
синапса центральной нервной
системы находят себе полное
объяснение при представлении о
гуморальной передаче возбуждения
с одного нейрона на другой.”
Казанский медицинский журнал,
1933.
13.
Классификации физиологииОпределение предмета
физиологии
14.
1. Физиология растений2. Физиология животных
3. Физиология человека
Классификации физиологии
Нормальная
Патологическая
Клиническая
15.
ФизиологияОбщая - основные жизненные процессы
Частная - свойства отдельных систем, тканей (сердечно-сосудистой и т.д.)
Прикладная - деятельность в связи со специфическими задачами и
условиями:
Физиология труда
Физиология спорта
Физиология питания
Экологическая физиология
Физиология сельскохозяйственных животных
Авиационная и космическая
Подводная
Возрастная – изучение функций организма в процессе развития
Сравнительная - клетки, органы, системы различных животных
Эволюционная- эволюция функций
Физиология адаптации - функционирование органов, тканей, систем во
взаимодействии с определенными природными факторами в различных
физико-географических зонах.
Физиология клетки
16.
Физиология- это наука о функциях ипроцессах, протекающих в
организме или составляющих его
органах, тканях, клетках и
механизмах их регуляции.
- о пределах нормы жизненных
процессов
Основной вопрос физиологии –
почему?
17. Методы физиологии 1. эксперимент (острый и хронический) 2. экстирпация (удаление) 3. трансплантация 4. фистульный 5.
катетеризация6. денервация
7. раздражение
8. изолированные органы, клетки
18. Физиология возбудимых тканей
19.
Все электрические процессыклетки разворачиваются на
плазматической (поверхностной )
мембране.
20. Клетка. Основные органеллы
21. Жидкостно-мозаичная модель плазматической мембраны
22.
Основные функции биологических мембран1. Граница между внутренней и наружной средами
клетки (отличаются физико-химическими показателями).
2. Участие во всех процессах обмена веществ
(ферментные системы, мембрана митохондрий и ЭПР).
3. Место, где протекает большинство биохимических
реакций.
4. Координируют и регулируют физиологические
процессы в клетках.
5. Образование межклеточных контактов
(обеспечивают их адгезию). Адгезия или сцепление
клеток друг с другом обеспечивает существование ткани.
23. Словарь
Возбудимость–
способность
живых
клеток
воспринимать раздражители и отвечать на
них
возбуждением.
Возбуждение – процесс перехода живой клетки из
состояния
покоя
в
состояние
активности,
сопровождается
возникновением
электрического
сигнала на мембране (потенциал действия, нервный
импульс).
Возбуждение запускает специфическую для каждой
ткани функцию. Возбуждение мембраны мышц
вызывает их сокращение, в нервной системе
возбуждение
мембраны
клетки
вызывает
его
проведение по аксону, в железистой ткани приводит к
секреции.
24.
Раздражитель - фактор внешней или внутреннейсреды, который оказывает влияние на возбудимую
клетку и изменяет ее активность
Порог
раздражения
интенсивность
раздражителя,
возбуждение. Чем ниже порог,
возбудимость
минимальная
вызывающая
тем больше
Раздражители классифицируются:
по адекватности (адекватные и неадекватные),
природе (физические, химические)
силе (подпороговые, пороговые и сверхпороговые).
25.
Все ткани организма в зависимостиот клеток, из которых они состоят,
делятся на:
- электровозбудимые,
- хемовозбудимые и
- механовозбудимые,
т.е. на те, которые под действием
того или иного раздражителя меняют
электрическое состояние мембраны.
26.
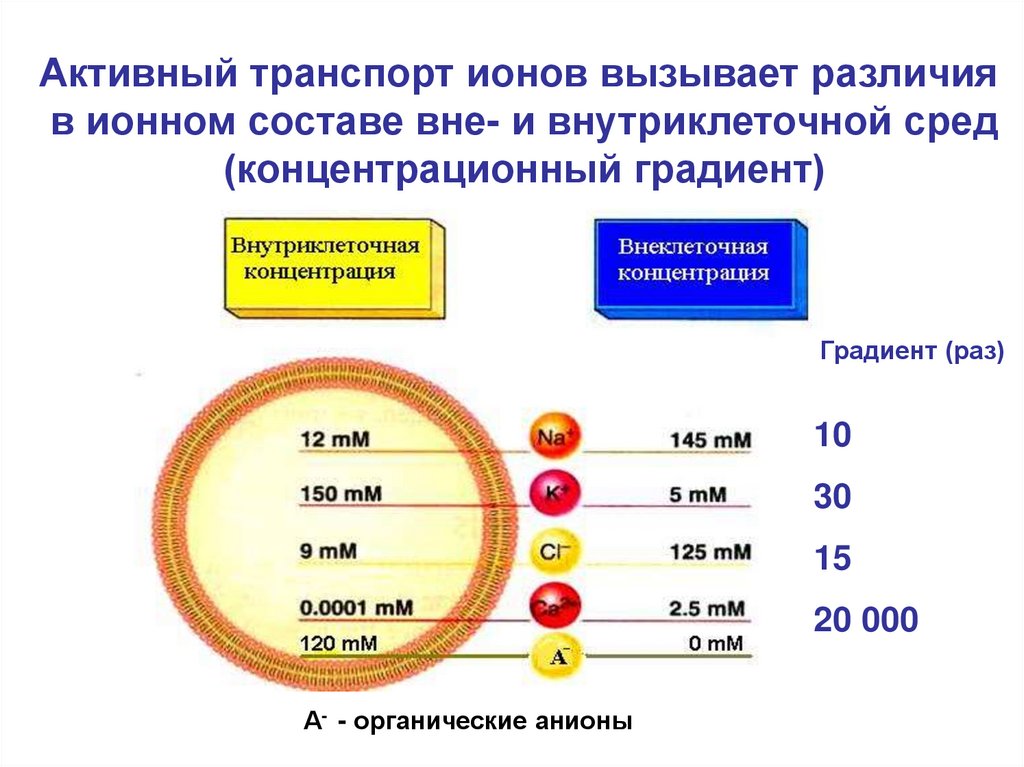
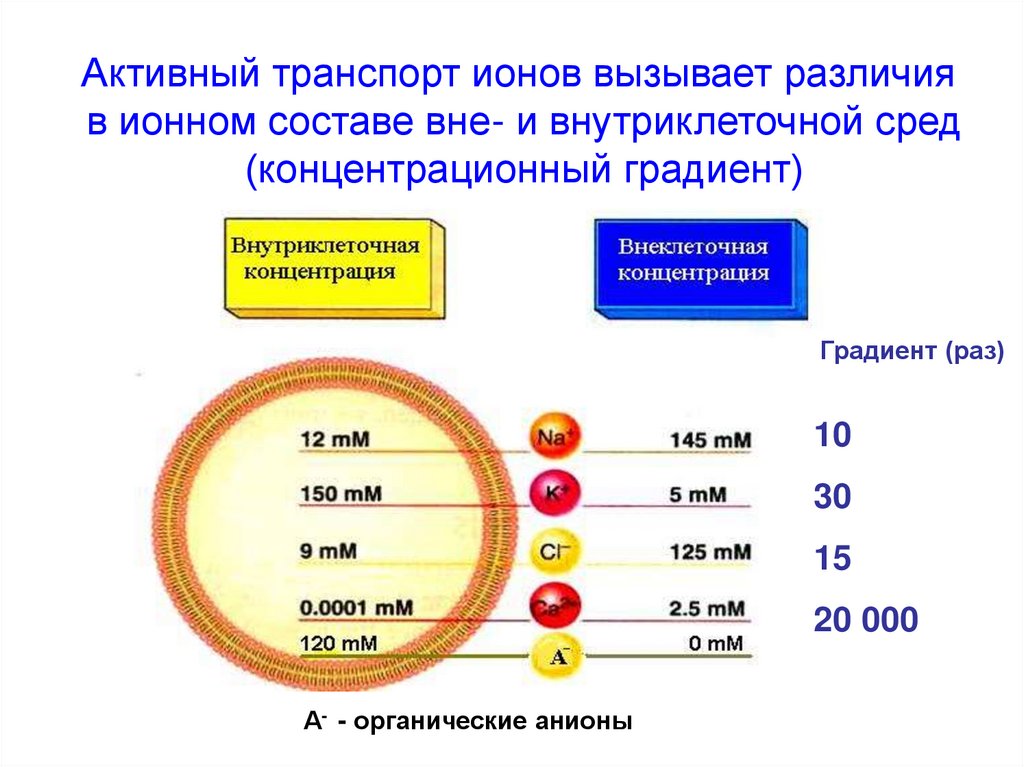
Активный транспорт ионов вызывает различияв ионном составе вне- и внутриклеточной сред
(концентрационный градиент)
Градиент (раз)
10
30
15
20 000
А- - органические анионы
27. Виды ионного транспорта
Активный - с затратой энергии АТФ,против концентрационного и/или
электрического градиента
• Первичный
• Вторичный
Пассивный – без затрат энергии, по
концентрационному и/или
электрическому градиенту
• Простая диффузия (ионные каналы)
• Облегченная диффузия (белки-переносчики)
• Осмос
28. Разность концентраций ионов снаружи и внутри клетки создается путем активного транспорта
Активный транспорт –первичный
(использование
Виды активного
транспорта
энергии
расщепления АТФ),
Направления транспорта ионов
Na+
Ca2+
K+
Ca2+
Cl-
вторичный (использование
энергии потока ионов по
градиенту концентрации).
Различают ко-транспорт
(движение ионов в одном
направлении) и ионообмен
(движение в противоположном
направлении)
29. Активный транспорт Натрий- калиевый обменный насос (АТФаза) Перенос ионов натрия и калия через мембрану. Принцип работы натрий-
Na+Активный транспорт
Натрий- калиевый обменный насос (АТФаза)
Перенос ионов натрия и калия через мембрану. Принцип работы
натрий- калиевого насоса (натрий- калиевой АТФазы)
K+
(А) Посадочные места, направленные внутрь
клетки, обладают высоким сродством к
натрию и низким — к калию. Ионы
калия, до этого момента связанные с
молекулой АТФазы, высвобождаются, в
то время как ионы натрия связываются с
ней.
(Б) Вслед за связыванием натрия происходит
связывание молекулы АТФ и
фосфорилирование фермента.
(В) В результате фосфорилирования в
структуре фермента происходят
изменения, приводящие к выдвижению
посадочных мест во внеклеточную
среду.
(Г) В таком положении посадочные места
обладают низким сродством к натрию и
высоким — к калию, поэтому ионы
натрия освобождаются во внеклеточную
среду, а ионы калия связываются с
молекулой АТФазы.
(Д) При связывании ионов калия АТФаза
дефосфорилируется.
(Е) Возврат в первоначальное состояние.
30. Активный транспорт Кальциевый насос (АТФаза). Натрий- кальциевый обменник
(А)Ca2+
Ca2+
(Б)
Кальциевый насос
(АТФаза)
(А) Внутриклеточная концентрация ионов кальция
поддерживается на очень низком уровне, за счет
первичного активного транспорта ионов кальция из
цитоплазмы
в
окружающую
среду
или
в
эндоплазматический
ретикулум
(в
мышце
саркоплазматический ретикулум).
Эффективность второго вида транспорта более
значительна, поскольку переносчик за один цикл
переносит 2 иона кальция.
Натрий- кальциевый
обменник
(Б) Натрий-кальциевый обменник.
Выброс одного иона кальция из
клетки энергетически сопряжен с
входом трех ионов натрия.
31. Активный транспорт Калий- хлорный ионообменник и натрий-калий-хлорный ко-транспорт
Cl -Cl Фуросемид-
(В)
блокатор транспорта хлора
1
Ионы2хлора могут как
закачиваться в клетку, так и
выкачиваться из нее.
(В) в некоторых нейронах
существует первичный активный
транспорт хлора наружу.
(Г) Калий-хлорный ко-транспорт
осуществляет вывод ионов хлора
из клетки за счет энергии
выходящего тока калия.
(Д) В некоторых нейронах и
мышечных волокнах транспорт
хлора внутрь клетки производится
натрий-калий-хлорным котранспортером .
1
(Г)
2
(Д)
32. Молекулярная структура транспортных молекул
(А) Натрий-калиевая АТФаза состоит из α-субъединицы, имеющей от 8 до 12трансмембранных спиральных сегментов, и меньшей по размеру β-субъединицы,
пронизывающей мембрану лишь один раз.
(Б) Натрий-кальциевые обменники имеют 11 трансмембранных сегментов.
(В) Калий-хлорные ко-транспортеры имеют 12 трансмембранных сегментов.
33. Транспорт ионов через ионные каналы
34. Модель облегченной диффузии глюкозы
35. Включение транспортных белков в состав мембраны
36. Модель работы активного транспорта
37.
Ионные каналы, строение, классификация,способ активации, прикладные аспекты
38.
Активный транспорт ионов вызывает различияв ионном составе вне- и внутриклеточной сред
(концентрационный градиент)
Градиент (раз)
10
30
15
20 000
А- - органические анионы
39. Зачем нужен концентрационный градиент?
• Электрические токи, возникающие вклетке, обеспечиваются пассивным
движением ионов через мембрану
• Для того, чтобы ионы могли двигаться
через мембрану, необходимо создать
разность концентраций снаружи и внутри
клетки (концентрационный градиент)
40. Виды ионного транспорта
• Активный - с затратой энергии АТФ, противконцентрационного и/или электрического
градиента
– Первичный
– Вторичный
• Пассивный – без затрат энергии, по
концентрационному и/или электрическому
градиенту
– Простая диффузия (ионные каналы)
– Облегченная диффузия (белки-переносчики)
– Осмос
41. Для того, чтобы ионы могли двигаться через мембрану, необходимо иметь мембранные структуры, сообщающие вне- и внутриклеточную
среду (ионные каналы)1-1000 каналов на квадратный микрометр мембраны
42. Как выглядит ионный канал?
• Центральная воднаяпора
• Устья канала
• Ворота
43. Два основных типа ионных каналов
В зависимости от роли в
нейрональной сигнализации,
различают 2 основных типа
ионных каналов – каналы покоя и
воротные -gate- (управляемые)
каналы.
Каналы покоя открываются в
покое без влияния внешних
факторов. Они участвуют,
преимущественно, в поддержании
мембранного потенциала покоя и
проницаемы для ионов К или Сl.
Большинство gate-каналов в
покое закрыто. Вероятность их
открытия регулируется
определенными воздействиями.
Они участвуют в генерации
электрических сигналов.
44. Работа канала
• Покой – канал закрыт, но может открытьсяпод действием адекватного стимула
• Активация- открытие канала под действием
адекватного стимула.
• Инактивация – состояние, когда канал закрыт
и адекватный стимул не действует (для
потенциалзависимых каналов) или
десенситизация –для лигандактивируемых
каналов)
45. Модель потенциал-зависимого ионного канала
Модельпотенциалзависимого
ионного
канала
46. Классификация ионных каналов
По избирательности1. Неселективные (никотиновый холинорецептор)
2. Селективные (Na+, K+, Ca++, Cl-)
По механизму активации
1.Потенциалзависимые (Na+, K+, Ca++)
2.Хемочувствительные (лигандактивируемые)
а) ионотропные рецепторы (Н-ХР, NMDA-Р, пуриновые Р)
б) активируются с цитоплазматической стороны Кса,
3.Механочувствительные (в волосковых клетках уха, в
кардиомиоцитах др.)
По проводимости
1. Большой проводимости Na+, К Са , АХ (рецептор)
2. Малой проводимости Ca++
47. Классификация ионных каналов
По скорости активациивероятность открытия)
(активация
увеличивает
1. Быстровозбудимые Na+
2. Медленно активируемые медленные К+ каналы
По инактивации
1. Инактивируемые Na+
2. Неинактивируемые медленные К+ каналы
По времени жизни
1. Короткоживущие (менее 1 мс ) Н-ХР, Na+
2. Долгоживущие (более 100 мс ) пуриновые рецепторы
48. Избирательность (селективность) каналов
• Селективные(Na+, К+, Са 2+, Cl- каналы ).
Селективность определяется
• размерами поры и иона,
• гидратной оболочкой,
• зарядом иона
• зарядом внутренней
поверхности канала
• Неселективные
49. Неселективный ионный канал Н-холинорецептор
50. Способы открытия (активации) управляемых ионных каналов
Активация физическимиизменениями
Потенциал-управляемые каналы
Каналы, активирующиеся
растяжением
Активация химическими
веществами
Внеклеточная активация
Внутриклеточная активация
51. Калиевые каналы
электровозбудимыебыстрые э/в
каналы
П-А-И
медленные
неинактивируе
мые
П-А
функция:
быстро вернуть Создание
мембранный
мембранного
потенциал к
потенциала
исходному
состоянию
после его
снижения
хемовозбудимые
М-ХР
АХ снаружи
К Са , К атф ,
Торможение
работы сердца
Предотвращает
перегрузку
сердца ионами
кальция
52. Кальциевые каналы
ПризнакПроводимость
l-каналы
(large)
T-каналы
(tiny)
N- каналы
(neuron)
P- каналы
(клетки
большая
малая
средняя
средняя
быстрая
средняя
быстрая
мышцы
сердце
периферичес- нейроны в
кие нейроны
головном
мозге
электромеханическое
сопряжение
возбуждение
секреция
медиатора
медленная
Скорость
активации и
инактивации
Объект
Функция
Пуркинье)
секреция
медиатора
53. Открытое и закрытое состояние ионных каналов
• Переход из закрытого в открытое состояниепроисходит моментально.
• Канал открывается на определенное время, которое
варьирует случайным образом. Среднее время
открытого состояния (мс).
• Активация - увеличение вероятности открытия канала
под действием адекватного стимула.
• Деактивация - снижение вероятности открытия канала
под действием адекватного стимула.
• Инактивация – переход канала в новое
конформационное состояние, когда адекватный
стимул не действует.
• Блокирование открытого состояния - токсины, ионы и
др.
54. Молекулярные механизмы активации и инактивации каналов
55. Проводимость и проницаемость каналов
• Величина тока, проходящего через канал, связана со скоростьюдвижения ионов через него и пропорциональна потенциалу на
мембране
i = gV,
где V –потенциал на мембране (в B),
i – величина тока через канал (в A),
константа g – проводимость канала (в Cм)
• Проводимость ионного канала зависит от легкости, с которой ионы
проходят через канал – проницаемости (внутреннее свойство
канала), и от концентрации ионов у устьев канала.
Ионный ток, текущий через мембрану клетки
I= i *P* N,
где i – ток через отдельный канал,
P- вероятность открытия канала,
N- количество каналов в мембране.
56. Строение ионного канала
Методыисследования
Аминокислоты
Выделение белков
каналов
Спиральные
сегменты
Аминокислотная
последовательность
Домены
Субъединицы
Клонирование
Канал
Точечные мутации
Экспрессия в
чужеродные клетки
Цитоплазма
57. Потенциал-управляемые селективные ионные каналы
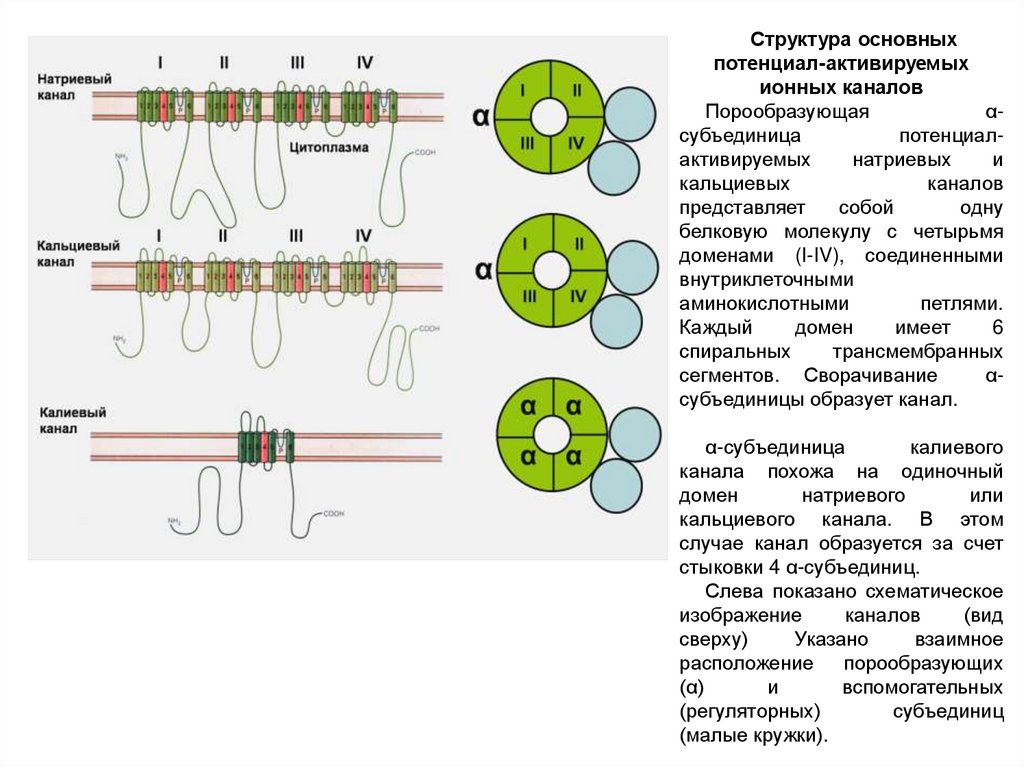
4 сенсора потенциала58.
Структура основныхпотенциал-активируемых
ионных каналов
Порообразующая
αсубъединица
потенциалактивируемых
натриевых
и
кальциевых
каналов
представляет
собой
одну
белковую молекулу с четырьмя
доменами (I-IV), соединенными
внутриклеточными
аминокислотными
петлями.
Каждый
домен
имеет
6
спиральных
трансмембранных
сегментов. Сворачивание
αсубъединицы образует канал.
α-субъединица
калиевого
канала похожа на одиночный
домен
натриевого
или
кальциевого канала. В этом
случае канал образуется за счет
стыковки 4 α-субъединиц.
Слева показано схематическое
изображение
каналов
(вид
сверху)
Указано
взаимное
расположение порообразующих
(α)
и
вспомогательных
(регуляторных)
субъединиц
(малые кружки).
59. Работа отдельного канала пэтч-кламп (patch-clamp)
Гигаомный контактПреимущества
1.
Возможность исследовать отдельный канал
2.
Возможность менять потенциал на мембране
3.
Возможность менять ионный состав и добавлять
любые исследуемые вещества с обоих сторон
мембраны
60. Нобелевская премия 1991 года в области физиологии и медицины
Эрвин Нейери
Берт Сакманн
«за открытия в области работы
одиночных ионных каналов»
61.
Что заставляет ионы двигатьсячерез открытые каналы?
62. Движение ионов через каналы
• Движение иона через каналуправляется двумя силами:
• 1) химической движущей
силой, которая зависит от
концентрационного градиента,
• 2) электрической движущей
силой, которая зависит от
разности электрического
потенциала на мембране.
• Потенциал на мембране, когда
электрическая сила точно
уравновешивается химической
силой и движение ионов через
канал прекращается назвали
равновесным потенциалом Е.
Е
цитоплазма
63. Равновесные потенциалы (Е) итоговая движущая сила (V- Е)
K-каналы-95
K+
Na-каналы
+67
Na+
Ca-каналы
+123
Ca++
Cl-каналы
-89 - 47 Cl-
i = gV,
i = g (V- Е)
64. Расчет равновесного потенциала
Равновесный потенциал для какого-либо иона Х
можно рассчитать из уравнения, полученного в 1888
году немецким физическим химиком Walter Nernst на
основании принципов термодинамики.
RT X o
ER
ln
zF X i
Где R – газовая постоянная, Т – температура (по
Келвину), z – валентность иона, F – константа
Фарадея, [Х]о и [Х]in – концентрации ионов внутри и
снаружи клетки.
• Уравнение Нернста можно использовать для расчета
равновесного потенциала любого иона по обе
стороны мембраны, проницаемой для данного иона.
65. Блокаторы ионных каналов
Прикладные медицинские аспектыБлокаторы ионных каналов
Na+ каналы
Тетродотоксин
(рыба фугу)
Сакситоксин
(планктон,
моллюски)
Лидокаин
Кокаин
Тетракаин
Прокаин
К+ каналы
Са2+ каналы
Тетраэтиламмоний Двухвалентные
4-аминопиридин
Ибериотоксин
(яд скорпиона)
катионы (кобальт,
никель, кадмий)
Дигидропиридины
(нитрендипин)
Фенилалкиламины
(верапамил)
Бензодиазепины
(дилтиазем)