









































 biology
biologySimilar presentations:










Введение в обмен веществ: биохимические основы регуляции метаболизма
1.
ЛЕКЦИЯВВЕДЕНИЕ В ОБМЕН ВЕЩЕСТВ:
БИОХИМИЧЕСКИЕ ОСНОВЫ РЕГУЛЯЦИИ
МЕТАБОЛИЗМА
Дисциплина: Б1.Б.8 –Биологическая химия, биохимия
полости рта
Специальность: 31.05.03 –Стоматология
НГМУ, КАФЕДРА МЕДИЦИНСКОЙ ХИМИИ
К.М.Н., ДОЦЕНТ ЛЕБЕРФАРБ ЕЛЕНА ЮРЬЕВНА
2.
АКТУАЛЬНОСТЬ ТЕМЫПостоянство гомеостаза в организме
обеспечивается несколькими регуляторными
системами (нервная, эндокринная, иммунная).
Все уровни регуляции в организме
интегрированы и действуют как единое целое.
3.
ПЛАН ЛЕКЦИИРоль гормонов в регуляции метаболизма
Механизмы действия гормонов
Регуляция обмена углеводов, липидов и
аминокислот
Основные этапы катаболизма
Энергетический обмен
Регуляция метаболизма основных
энергетических субстратов
4.
БИОХИМИЧЕСКИЕ ОСНОВЫ ГОРМОНАЛЬНОЙРЕГУЛЯЦИИ ОБМЕНА ВЕЩЕСТВ
Эндокринная система обеспечивает регуляцию и
интеграцию метаболизма в разных тканях в ответ на
изменения условий внешней и внутренней среды.
Гормоны функционируют как химические
посредники, переносящие информацию об этих
изменениях в различные органы и ткани.
Ответная реакция клетки на действие гормона
определяется как химическим строением гормона,
так и типом клетки, на которую направлено его
действие.
Гормоны присутствуют в крови в очень низкой
концентрации, и их действие обычно
кратковременно.
5.
Гормоны (греч. hormao – привожу в движение) – это вещества, кпризнакам которых относятся:
синтез в специализированных клетках,
регулирование обмена веществ в отдельных органах и во всем
организме в целом,
высокая специфичность действия (влияние на конкретные
органы-мишени либо на определенные метаболические процессы),
высокая биологическая активность в очень малых количествах,
большая длительность действия (по сравнению с нервной
системой),
дистантный характер действия (удаленность мишени от места
синтеза гормона),
действуют через рецепторы, изменяя активность ферментативных
систем.
С нарушением гормонального обмена связан ряд наследственных и
приобретенных заболеваний, сопровождающихся серьезными
проблемами в развитии и жизнедеятельности организма
(карликовость и гигантизм, сахарный и несахарный диабет,
микседема, бронзовая болезнь и др).
6.
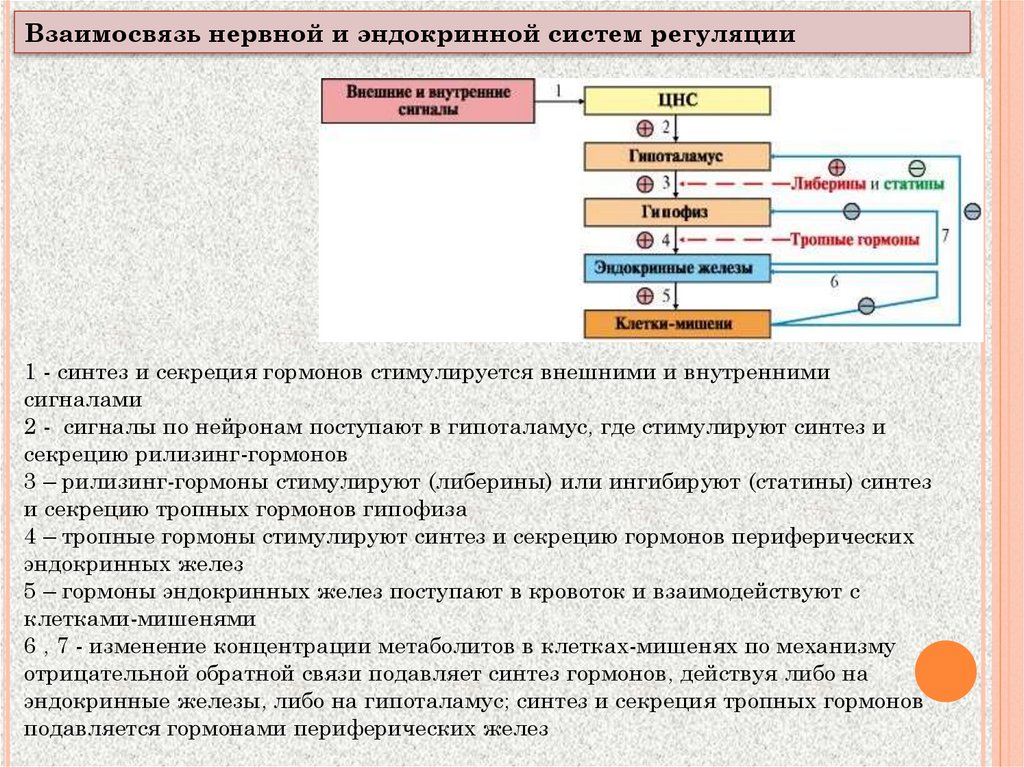
Взаимосвязь нервной и эндокринной систем регуляции1 - синтез и секреция гормонов стимулируется внешними и внутренними
сигналами
2 - сигналы по нейронам поступают в гипоталамус, где стимулируют синтез и
секрецию рилизинг-гормонов
3 – рилизинг-гормоны стимулируют (либерины) или ингибируют (статины) синтез
и секрецию тропных гормонов гипофиза
4 – тропные гормоны стимулируют синтез и секрецию гормонов периферических
эндокринных желез
5 – гормоны эндокринных желез поступают в кровоток и взаимодействуют с
клетками-мишенями
6 , 7 - изменение концентрации метаболитов в клетках-мишенях по механизму
отрицательной обратной связи подавляет синтез гормонов, действуя либо на
эндокринные железы, либо на гипоталамус; синтез и секреция тропных гормонов
подавляется гормонами периферических желез
7.
КЛАССИФИКАЦИЯ ПО СТРОЕНИЮГормоны – производные аминокислот
(адреналин, норадреналин, тироксин,
трийодтиронин)
Белково-пептидные гормоны (тропные гормоны
гипофиза (СТГ, АКТГ, ТТГ,МСГ, ФСГ, ЛГ),
инсулин, глюкагон, АДГ, кальцитонин,
паратгормон)
Стероидные гормоны (кортизол, кальцитриол,
альдостерон, тестостерон, эстрадиол)
8.
МЕХАНИЗМЫ ПЕРЕДАЧИ ГОРМОНАЛЬНЫХ СИГНАЛОВ ВКЛЕТКИ
9.
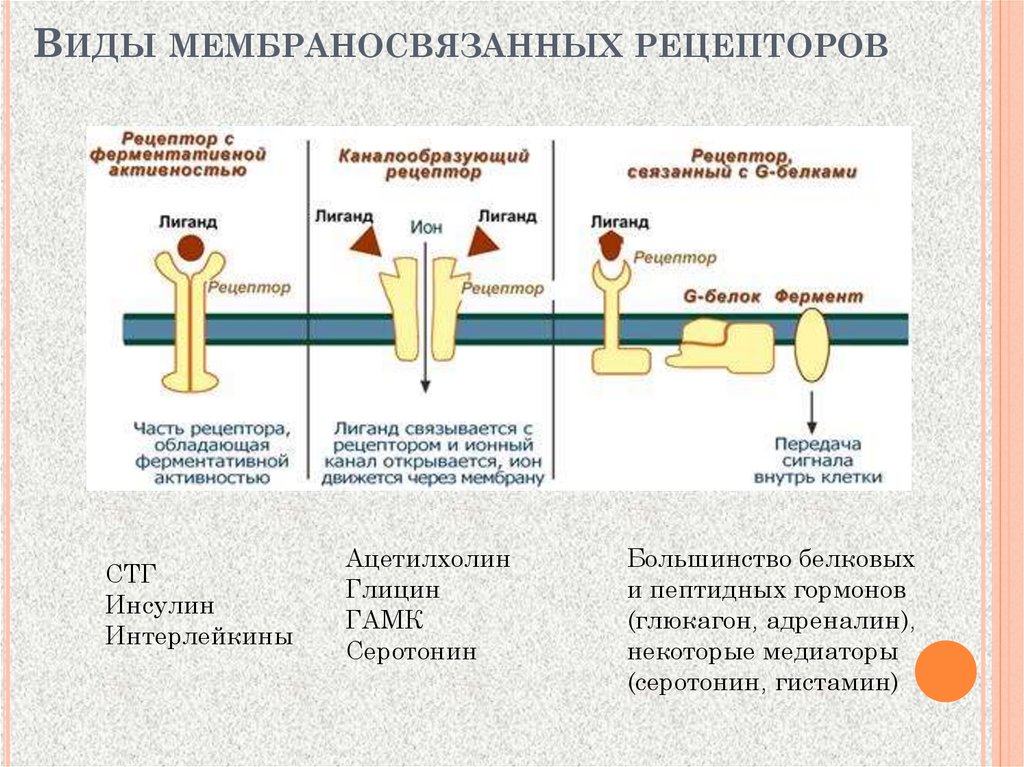
ВИДЫ МЕМБРАНОСВЯЗАННЫХ РЕЦЕПТОРОВСТГ
Инсулин
Интерлейкины
Ацетилхолин
Глицин
ГАМК
Серотонин
Большинство белковых
и пептидных гормонов
(глюкагон, адреналин),
некоторые медиаторы
(серотонин, гистамин)
10.
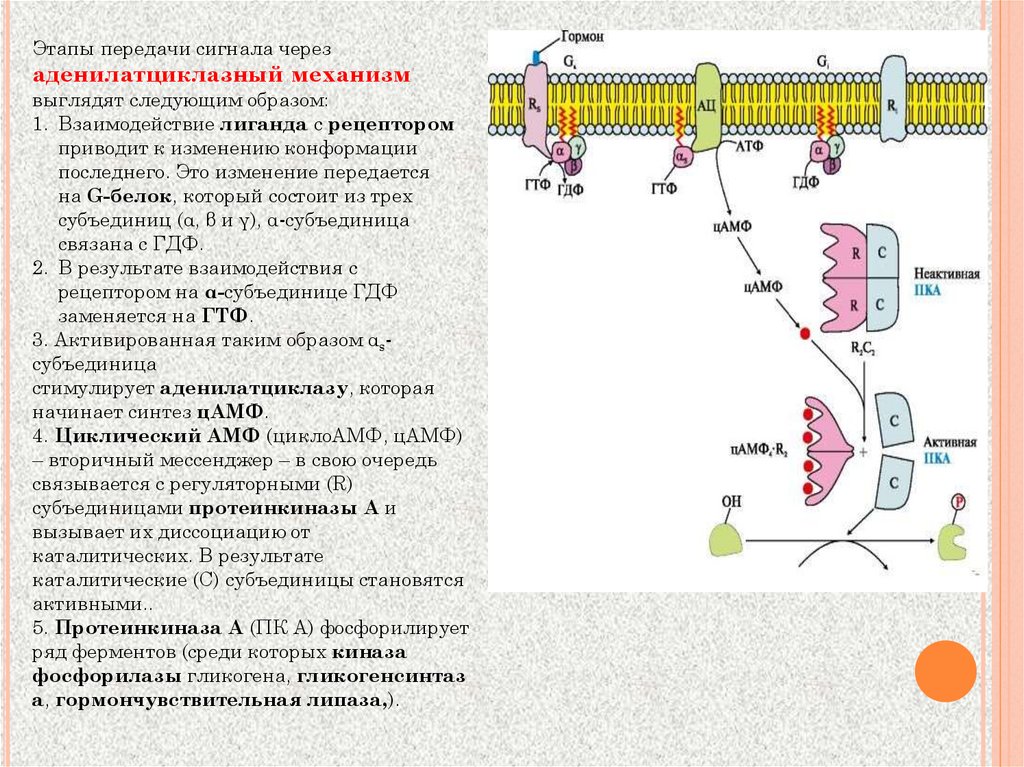
Этапы передачи сигнала черезаденилатциклазный механизм
выглядят следующим образом:
1. Взаимодействие лиганда с рецептором
приводит к изменению конформации
последнего. Это изменение передается
на G-белок, который состоит из трех
субъединиц (α, β и γ), α-субъединица
связана с ГДФ.
2. В результате взаимодействия с
рецептором на α-субъединице ГДФ
заменяется на ГТФ.
3. Активированная таким образом αsсубъединица
стимулирует аденилатциклазу, которая
начинает синтез цАМФ.
4. Циклический АМФ (циклоАМФ, цАМФ)
– вторичный мессенджер – в свою очередь
связывается с регуляторными (R)
субъединицами протеинкиназы А и
вызывает их диссоциацию от
каталитических. В результате
каталитические (C) субъединицы становятся
активными..
5. Протеинкиназа А (ПК А) фосфорилирует
ряд ферментов (среди которых киназа
фосфорилазы гликогена, гликогенсинтаз
а, гормончувствительная липаза,).
11.
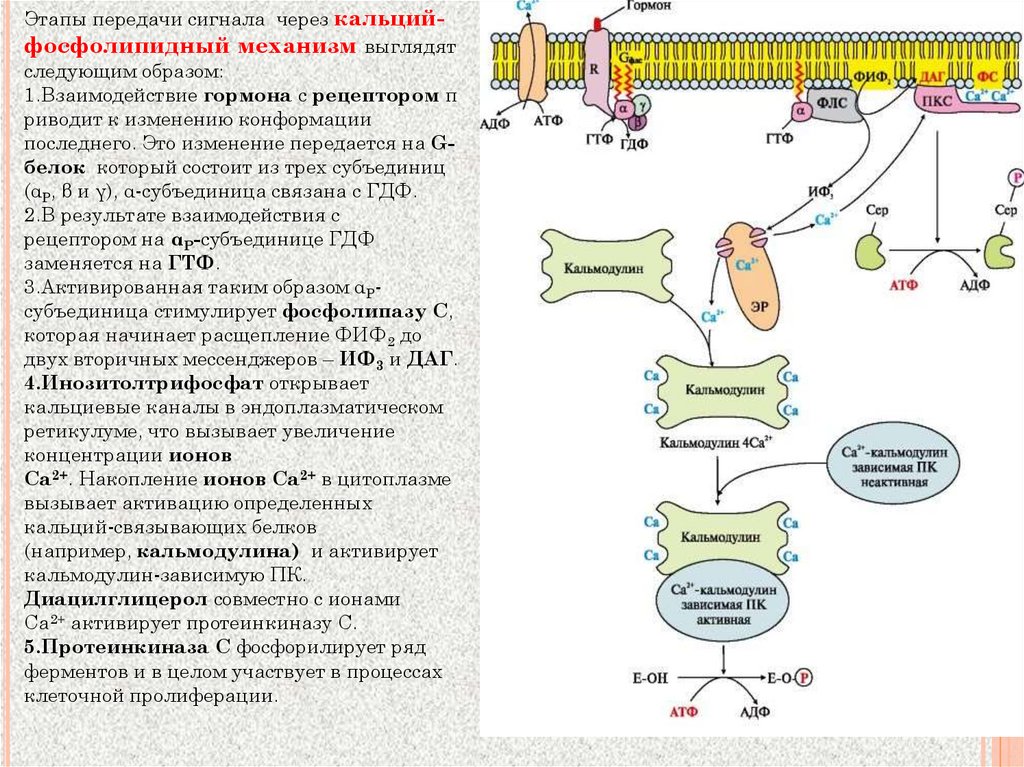
Этапы передачи сигнала через кальцийфосфолипидный механизм выглядятследующим образом:
1.Взаимодействие гормона с рецептором п
риводит к изменению конформации
последнего. Это изменение передается на Gбелок который состоит из трех субъединиц
(αP, β и γ), α-субъединица связана с ГДФ.
2.В результате взаимодействия с
рецептором на αP-субъединице ГДФ
заменяется на ГТФ.
3.Активированная таким образом αPсубъединица стимулирует фосфолипазу С,
которая начинает расщепление ФИФ2 до
двух вторичных мессенджеров – ИФ3 и ДАГ.
4.Инозитолтрифосфат открывает
кальциевые каналы в эндоплазматическом
ретикулуме, что вызывает увеличение
концентрации ионов
Cа2+. Накопление ионов Са2+ в цитоплазме
вызывает активацию определенных
кальций-связывающих белков
(например, кальмодулина) и активирует
кальмодулин-зависимую ПК.
Диацилглицерол совместно с ионами
Са2+ активирует протеинкиназу С.
5.Протеинкиназа С фосфорилирует ряд
ферментов и в целом участвует в процессах
клеточной пролиферации.
12.
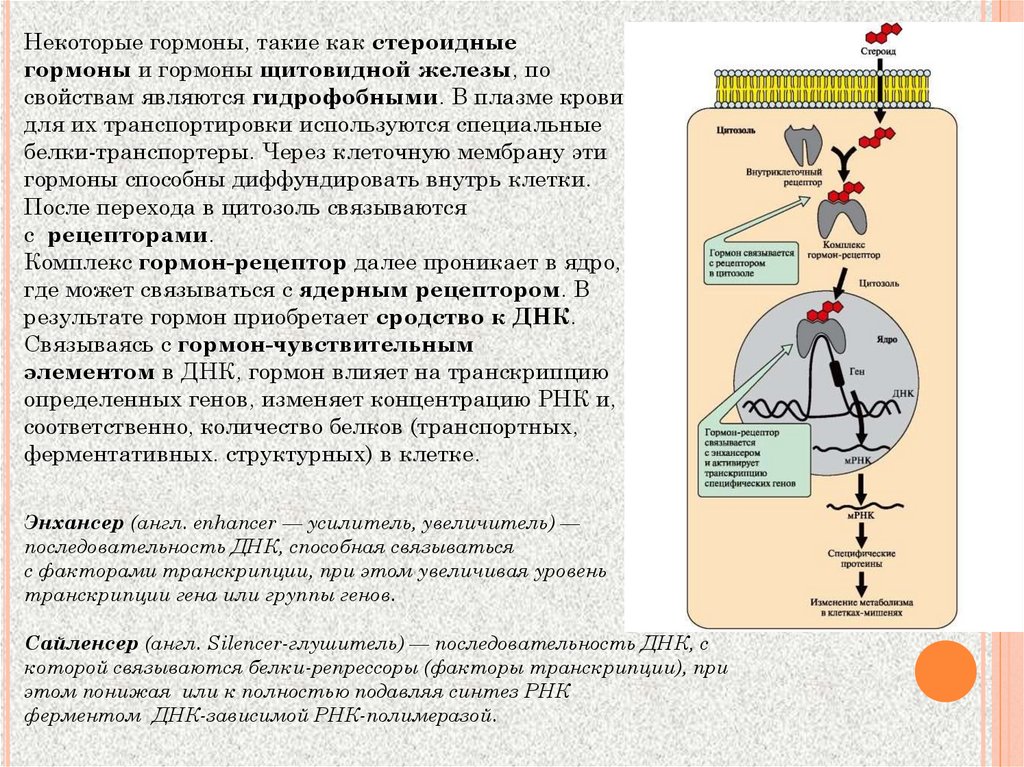
Некоторые гормоны, такие как стероидныегормоны и гормоны щитовидной железы, по
свойствам являются гидрофобными. В плазме крови
для их транспортировки используются специальные
белки-транспортеры. Через клеточную мембрану эти
гормоны способны диффундировать внутрь клетки.
После перехода в цитозоль связываются
с рецепторами.
Комплекс гормон-рецептор далее проникает в ядро,
где может связываться с ядерным рецептором. В
результате гормон приобретает сродство к ДНК.
Связываясь с гормон-чувствительным
элементом в ДНК, гормон влияет на транскрипцию
определенных генов, изменяет концентрацию РНК и,
соответственно, количество белков (транспортных,
ферментативных. структурных) в клетке.
Энхансер (англ. enhancer — усилитель, увеличитель) —
последовательность ДНК, способная связываться
с факторами транскрипции, при этом увеличивая уровень
транскрипции гена или группы генов.
Сайленсер (англ. Silencer-глушитель) — последовательность ДНК, с
которой связываются белки-репрессоры (факторы транскрипции), при
этом понижая или к полностью подавляя синтез РНК
ферментом ДНК-зависимой РНК-полимеразой.
13.
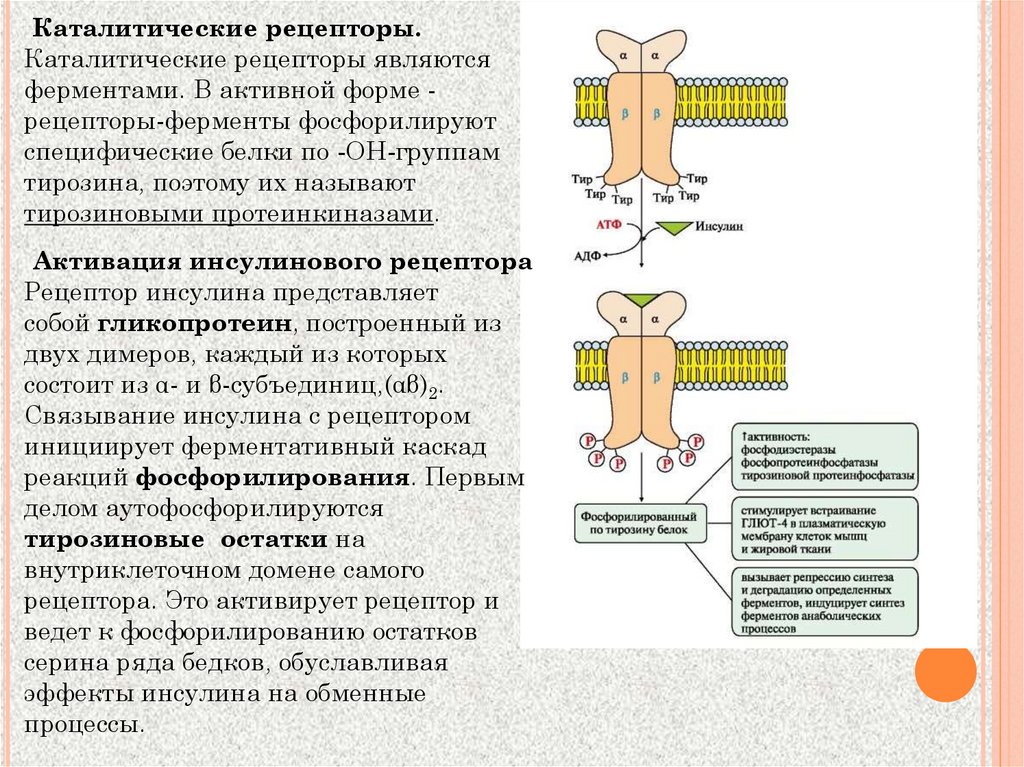
Каталитические рецепторы.Каталитические рецепторы являются
ферментами. В активной форме рецепторы-ферменты фосфорилируют
специфические белки по -ОН-группам
тирозина, поэтому их называют
тирозиновыми протеинкиназами.
Активация инсулинового рецептора
Рецептор инсулина представляет
собой гликопротеин, построенный из
двух димеров, каждый из которых
состоит из α- и β-субъединиц,(αβ)2.
Связывание инсулина с рецептором
инициирует ферментативный каскад
реакций фосфорилирования. Первым
делом аутофосфорилируются
тирозиновые остатки на
внутриклеточном домене самого
рецептора. Это активирует рецептор и
ведет к фосфорилированию остатков
серина ряда бедков, обуславливая
эффекты инсулина на обменные
процессы.
14.
ПЛАН ХАРАКТЕРИСТИКИ ГОРМОНОВРЕГУЛЯТОРОВ МЕТАБОЛИЧЕСКИХ ПРОЦЕССОВХимическая природа гормона (белково-пептидный,
стероидный, производное аминокислоты)
Место синтеза (органы, ткани)
Особенности синтеза (для стероидных и тиреоидных гормонов,
адреналина)
Сигнал для секреции гормона
Транспортная форма гормона в крови (для стероидных и
тиреоидных гормонов)
Мишени гормона (органы, ткани)
Механизм действия гормона (аденилатциклазный,
фосфолипазный, геномный; ключевые ферменты, активность
которых гормон повышает и запускает таким образом
соответствующие метаболические процессы)
Конечный биологический эффект
14
15.
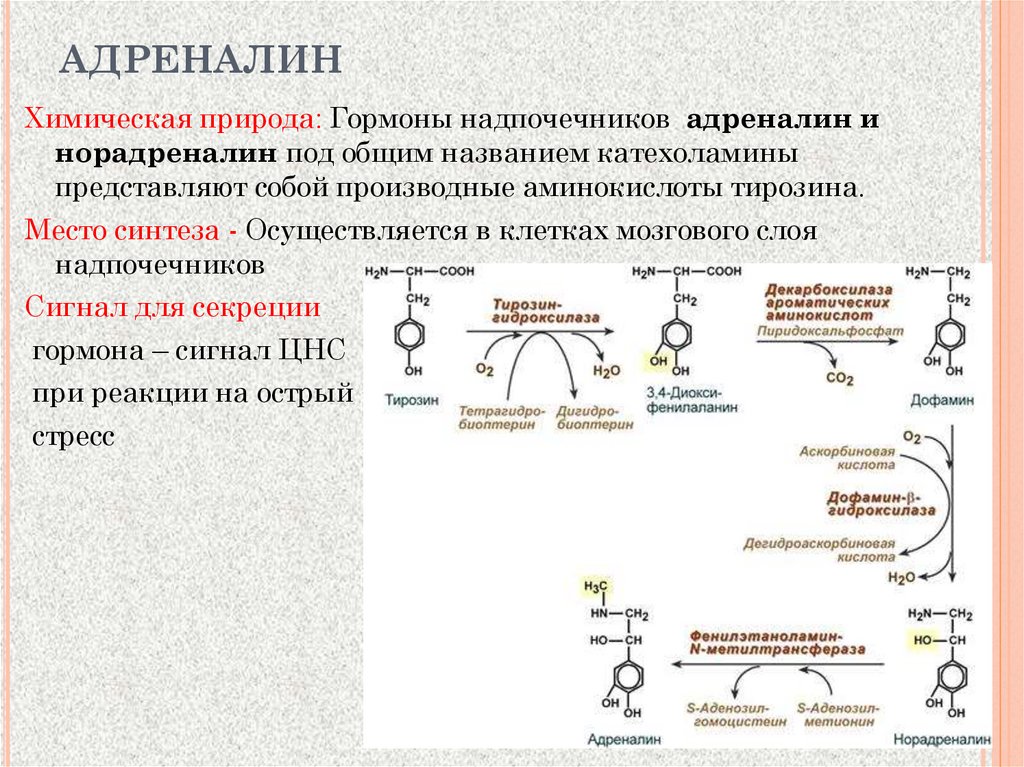
АДРЕНАЛИНХимическая природа: Гормоны надпочечников адреналин и
норадреналин под общим названием катехоламины
представляют собой производные аминокислоты тирозина.
Место синтеза - Осуществляется в клетках мозгового слоя
надпочечников
Сигнал для секреции
гормона – сигнал ЦНС
при реакции на острый
стресс
16.
Механизм действия гормона - Механизм действия гормонов разный взависимости от рецептора.
Например, в жировой ткани при низких концентрациях адреналина более
активны α2-адренорецепторы, при повышенных концентрациях (стресс)
– стимулируются β1-, β2-, β3-адренорецепторы.
при возбуждении α1-адренорецепторов – инозитолфосфатный
при задействовании β1- и β2-адренорецепторов аденилатциклазный
Мишени гормона (органы, ткани) и биологический эффект
Печень - Активация гликогенолиза - повышение концентрации
глюкозы в крови
Мышцы – стимуляция гликогенолиза в скелетных мышцах;
Усиление сократительной активности скелетных мышц
расслабление гладких мышц трахеи, бронхов, кишечника
Сокращение гладких мышц кровеносных сосудов в разных областях
тела
Жировая ткань – стимуляция липолиза в жировой ткани
17.
ГЛЮКАГОНХимическая природа гормона - пептидный гормон, состоит из
29 аминокислот
Место синтеза – предшественник гормона синтезируется αклетками поджелудочной железы и превращается в глюкагон
путем частичного протеолиза
Сигнал для секреции – снижение уровня глюкозы в крови,
«гормон голода»
Мишени – печень и жировая ткань
Механизм действия гормона – аденилатциклазный (в жировой
ткани активирует ТАГ-липазу; в печени активирует
гликогенфосфорилазу и ферменты глюконеогенеза)
биологический эффект – повышение концентрации глюкозы
и жирных кислот в крови
18.
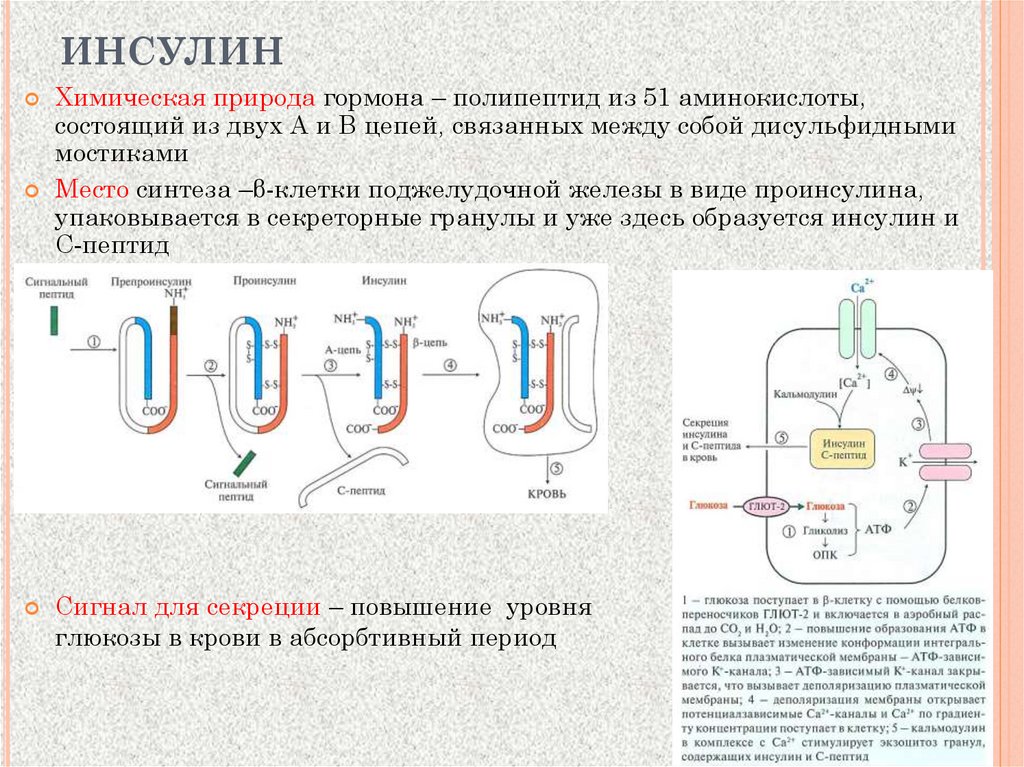
ИНСУЛИНХимическая природа гормона – полипептид из 51 аминокислоты,
состоящий из двух А и В цепей, связанных между собой дисульфидными
мостиками
Место синтеза –β-клетки поджелудочной железы в виде проинсулина,
упаковывается в секреторные гранулы и уже здесь образуется инсулин и
С-пептид
Сигнал для секреции – повышение уровня
глюкозы в крови в абсорбтивный период
19.
МЕХАНИЗМ ДЕЙСТВИЯ ГОРМОНАБыстрые эффекты
Быстрые эффекты заключаются в изменении скоростей
фосфорилирования и дефосфорилирования метаболических
ферментов и регуляторных белков.
Печень
торможение эффектов адреналина и глюкагона (фосфодиэстераза),
ускорение гликогеногенеза (гликогенсинтаза),
активация гликолиза (фосфофруктокиназа, пируваткиназа),
превращение пирувата в ацетил-SКоА (ПВК-дегидрогеназа),
усиление синтеза жирных кислот (ацетил-SКоА-карбоксилаза),
формирование ЛПОНП,
повышение синтеза холестерина (ГМГ-SКоА-редуктаза),
20.
Мышцыторможение эффектов адреналина (фосфодиэстераза),
стимулирует транспорт глюкозы в клетки (активация ГлюТ-4),
стимуляция гликогеногенеза (гликогенсинтаза),
активация гликолиза (фосфофруктокиназа, пируваткиназа),
превращение пирувата в ацетил-SКоА (ПВК-дегидрогеназа),
усиливает транспорт нейтральных аминокислот в мышцы,
стимулирует трансляцию (рибосомальный синтез белков).
Жировая ткань
стимулирует транспорт глюкозы в клетки (активация Глют-4),
активирует запасание жирных кислот в тканях
(липопротеинлипаза),
активация гликолиза (фосфофруктокиназа, пируваткиназа),
усиление синтеза жирных кислот (активация ацетил-SКоАкарбоксилазы),
создание возможности для запасания ТАГ (инактивация
гормон-чувствительной-липазы).
21.
Медленные эффекты (минуты-часы)Медленные эффекты заключаются в изменении
скорости транскрипции генов белков, отвечающих за обмен веществ, за
рост и деление клеток, например:
1. Индукция синтеза ферментов в печени глюкокиназы и пируваткиназы
(гликолиз),АТФ-цитрат-лиазы, ацетил-SКоА-карбоксилазы, синтазы
жирных кислот(синтез жирных кислот),глюкозо-6фосфатдегидрогеназы (пентозофосфатный путь),
2. Индукция в адипоцитах синтеза глицеральдегидфосфат-дегидрогеназы
и синтазы жирных кислот.
3. Репрессия синтеза мРНК, например, для ФЕП-карбоксикиназы
(глюконеогенез)
КОНЕЧНЫЙ БИОЛОГИЧЕСКИЙ ЭФФЕКТ
Действие инсулина направлено на ускорение анаболических
процессов в абсорбтивный период: увеличение поступления
глюкозы, жирных кислот и аминокислот в ткани, синтеза гликогена
в печени и мышцах, синтеза жирных кислот и ТАГ в печени и
жировой ткани, синтеза белков во многих тканях.
22.
КОРТИЗОЛХимическая природа – гормон
стероидной природы, производное
холестерола
Место синтеза – сетчатая и пучковая
зоны коры надпочечников
Синтез и секреция кортизола
регулируются гипоталамогипофизарной системой и
стимулируются при различных видах
продолжительного стресса.
Регулируется синтез по принципу
отрицательной обратной связи.
Транспортный белок – транскортин
Мишенью
является лимфоидная, эпителиаль
ная (слизистые оболочки и
кожа), жировая, костная и мышеч
ная ткани, печень.
Механизм действия - цитозольный
23.
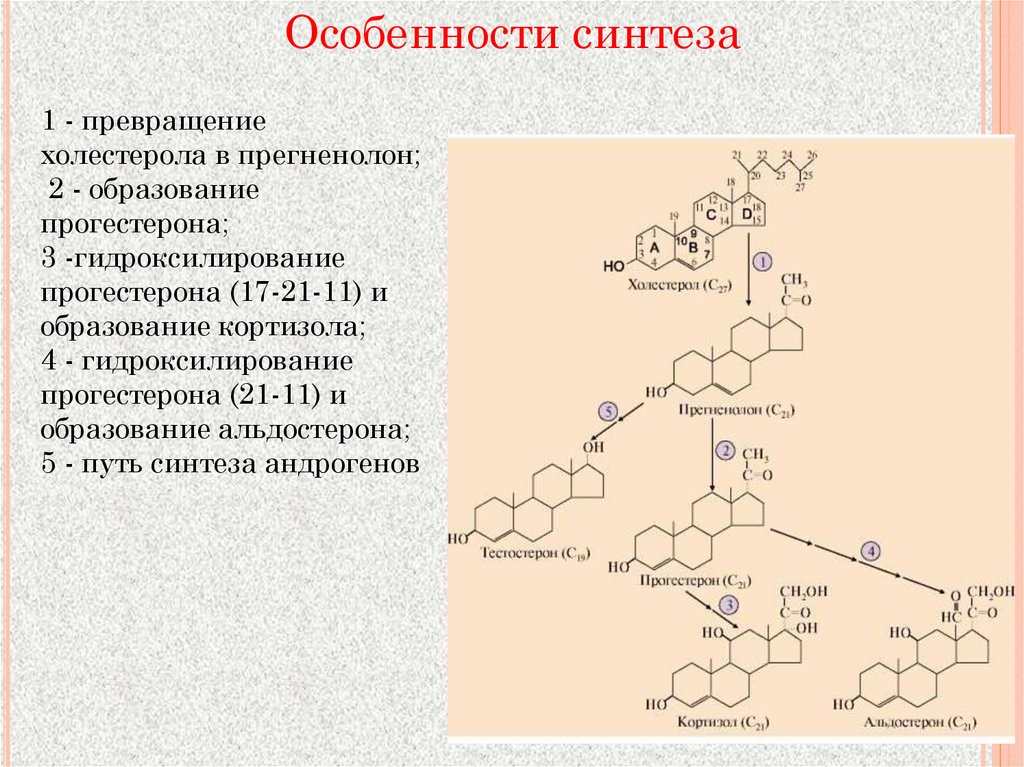
Особенности синтеза1 - превращение
холестерола в прегненолон;
2 - образование
прогестерона;
3 -гидроксилирование
прогестерона (17-21-11) и
образование кортизола;
4 - гидроксилирование
прогестерона (21-11) и
образование альдостерона;
5 - путь синтеза андрогенов
24.
БИОЛОГИЧЕСКИЙ ЭФФЕКТБелковый обмен
значительное повышение катаболизма белков в лимфоидной,
эпителиальной, мышечной, соединительной и костной тканях,
в печени в целом стимулирует анаболизм белков (например,
ферменты трансаминирования и глюконеогенеза),
стимуляция реакций трансаминирования через синтез
аминотрансфераз, обеспечивающих удаление аминогрупп от
аминокислот и получение углеродного скелета кетокислот,
Углеводный обмен
В целом вызывают повышение концентрации глюкозы крови:
усиление мощности глюконеогенеза из кетокислот за счет
увеличения синтеза фосфоенолпируват-карбоксикиназы,
увеличение синтеза гликогена в печени за счет активации
фосфатаз и дефосфорилирования гликогенсинтазы.
снижение проницаемости мембран для глюкозы в
инсулинзависимых тканях.
25.
Липидный обменстимуляция липолиза в жировой ткани благодаря увеличению
синтеза ТАГ-липазы
Водно-электролитный обмен
слабый минералокортикоидный эффект на канальцы почек
вызывает реабсорбцию натрия и потерю калия,
Противовоспалительное и иммунодепрессивное действие
увеличение перемещения лимфоцитов, моноцитов, эозинофилов
и базофилов в лимфоидную ткань,
повышение уровня лейкоцитов в крови за счет их выброса из
костного мозга и тканей,
подавление функций лейкоцитов и тканевых макрофагов через
снижение синтеза эйкозаноидов посредством уменьшения
транскрипции ферментов фосфолипазы А2 и циклооксигеназы.
26.
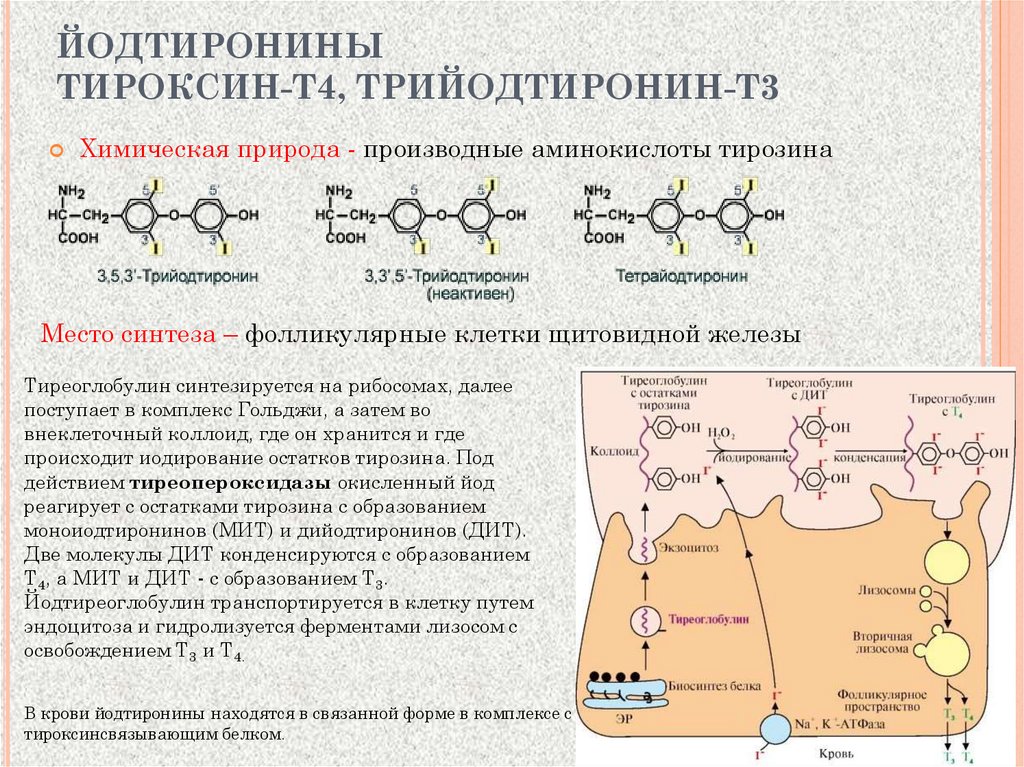
ЙОДТИРОНИНЫТИРОКСИН-Т4, ТРИЙОДТИРОНИН-Т3
Химическая природа - производные аминокислоты тирозина
Место синтеза – фолликулярные клетки щитовидной железы
Тиреоглобулин синтезируется на рибосомах, далее
поступает в комплекс Гольджи, а затем во
внеклеточный коллоид, где он хранится и где
происходит иодирование остатков тирозина. Под
действием тиреопероксидазы окисленный йод
реагирует с остатками тирозина с образованием
моноиодтиронинов (МИТ) и дийодтиронинов (ДИТ).
Две молекулы ДИТ конденсируются с образованием
Т4, а МИТ и ДИТ - с образованием Т3.
Йодтиреоглобулин транспортируется в клетку путем
эндоцитоза и гидролизуется ферментами лизосом с
освобождением Т3 и Т4.
В крови йодтиронины находятся в связанной форме в комплексе с
тироксинсвязывающим белком.
27.
Регуляция синтеза и секрецииАктивируют: тиреотропин на этапах поглощения йода, синтеза
тиреоглобулина, эндоцитоза и секреции Т3 и Т4 в кровь.
Уменьшают: тироксин и трийодтиронин (по механизму обратной
отрицательной связи).
Мишени гормона - ВСЕ
ткани организма
Механизм действия гормона
- цитозольный
Тиреоидные гормоны увеличивают
скорость базального метаболизма.
Главным эффектом является повышение
активности Na+,K+-АТФазы, что приводит
к быстрому расходованию АТФ и по
механизму дыхательного контроля
запускает катаболизм углеводов и
липидов. В митохондриях увеличивается
количество АТФ/АДФ-транслоказы и
потребление кислорода.
Сопутствующим эффектом усиления
катаболизма является термогенез
(наработка тепла).
28.
Белковый обмен: Усиливает транспорт аминокислот в клетки.Активирует синтез дифференцировочных белков в ЦНС, гонадах, костной
ткани и обусловливает развитие этих тканей.
У детей действие тиреоидных гормонов в целом анаболическое, т.к.
трийодтиронин усиливает выделение соматолиберина, что стимулирует
секрецию гормона роста.
У взрослых действие тиреоидных гормонов, в основном,
катаболическое.
Углеводный обмен: Увеличивает гликогенолиз и аэробное окисление
глюкозы.
Липидный обмен: Стимулирует липолиз, β-окисление жирных кислот,
подавляет стероидогенез.
Нуклеиновый обмен: Активирует начальные стадии синтеза
пуринов и синтеза пиримидинов, стимулирует дифференцировочный
синтез РНК и ДНК.
29.
ЗАДАНИЕ ДЛЯ САМОСТОЯТЕЛЬНОЙРАБОТЫ
Используя материал слайда №15 и
материал учебника составьте
характеристику тканевых гормонов,
производных ВЖК (эйкозаноидов)
Объясните принцип лекарственной
регуляции синтеза эйкозаноидов на
примере нестероидных
противовоспалительных препаратов
(аспирин, ибупрофен) и препаратов
кортизола (преднизолон,
дексаметазон)
30.
ЭНЕРГЕТИЧЕСКИЙ ОБМЕНМетаболизм, или обмен веществ - это набор
химических реакций, которые протекают в живом
организме обеспечивая его жизнедеятельность.
включает два неразрывных процесса анаболизм и
катаболизм.
Функции метаболизма:
Энергетическая – снабжение клетки химической
энергией,
Пластическая – синтез макромолекул,
Специфическая – синтез и распад биомолекул,
необходимых для выполнения специфических
клеточных функций.
31.
АНАБОЛИЗМАнаболизм – это биосинтез сложных органических
макромолекул из молекул-предшественников.
Поскольку он сопровождается усложнением
структуры, то требует затрат энергии. Источником
такой энергии является энергия АТФ.
32.
КАТАБОЛИЗМКатаболизм – расщепление и окисление сложных
органических молекул до более простых конечных
продуктов. Реакции катаболизма сопровождаются
высвобождением энергии, заключенной в сложной
структуре веществ. Часть высвобожденной энергии
рассеивается в виде тепла. Часть этой энергии
используется для синтеза АТФ.
33.
В высокоэргических связях АТФаккумулируется энергия, выделяемая в
процессах катаболизма. Энергия,
выделяющаяся при окислении
пищевых веществ, обеспечивает синтез
АТФ из АДФ и Н3РО4, а энергия
гидролиза АТФ, в свою очередь,
используется в различных видах
работы
Два способа синтеза АТФ
1. Основным способом получения АТФ в клетке является окислительное
фосфорилирование, протекающее в структурах внутренней мембраны митохондрий.
При этом энергия атомов водорода молекул НАДН и ФАДН2, образованных в
гликолизе и ЦТК, при окислении жирных кислот и аминокислот, преобразуется в
энергию связей АТФ.
2. Субстратное фосфорилирование. Этот способ связан с передачей макроэргического
фосфата или энергии макроэргической связи какого-либо вещества (субстрата) на
АДФ. К таким веществам относятся метаболиты гликолиза (1,3-дифосфоглицериновая
кислота, фосфоенолпируват), цикла трикарбоновых кислот (сукцинил-SКоА) и
резервный макроэрг креатинфосфат.
34.
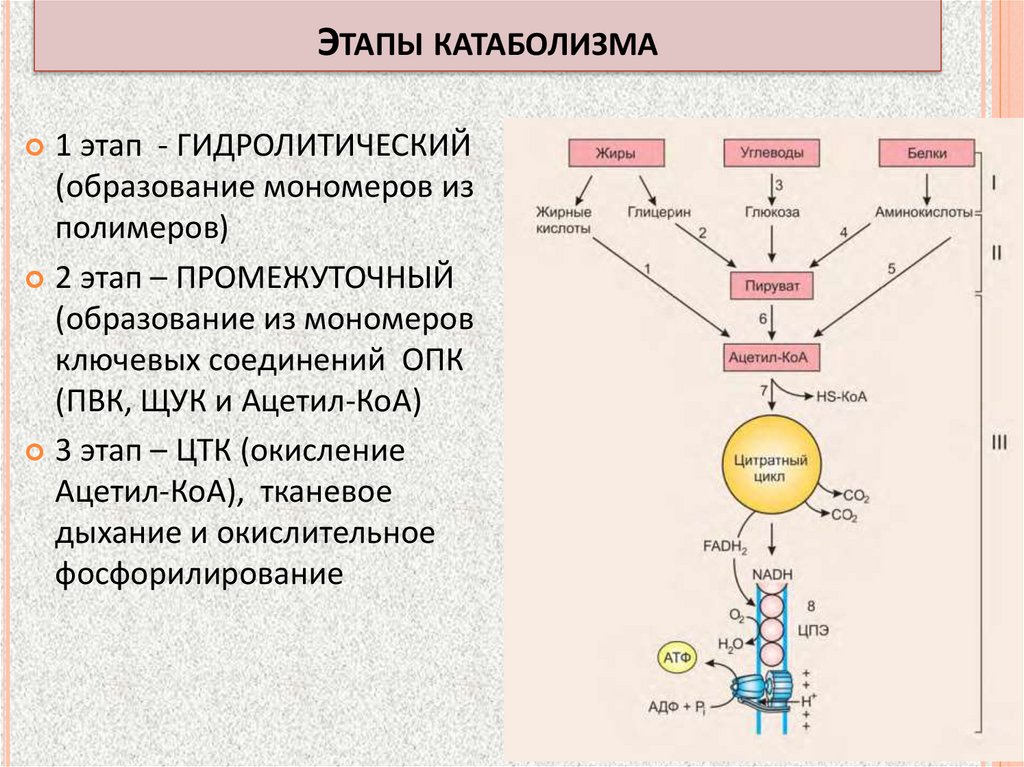
ЭТАПЫ КАТАБОЛИЗМА1 этап - ГИДРОЛИТИЧЕСКИЙ
(образование мономеров из
полимеров)
2 этап – ПРОМЕЖУТОЧНЫЙ
(образование из мономеров
ключевых соединений ОПК
(ПВК, ЩУК и Ацетил-КоА)
3 этап – ЦТК (окисление
Ацетил-КоА), тканевое
дыхание и окислительное
фосфорилирование
35.
Первый этап - происходит в жкт (перевариваниепищи) или в лизосомах (самообновление клеток).
При этом освобождается около 1% энергии, заключенной в
молекуле. Она рассеивается в виде тепла.
Второй этап – происходит в цитозоле и митохондриях.
При этом вещества, образованные в первом этапе
превращаются
в ПВК (моносахариды в гликолизе)
в ПВК, в Ацетил-КоА (катаболизма аминокислот)
в Ацетил-КоА (при β- окислении жирных кислот)
На этом этапе выделяется около 30% энергии, заключенной в
молекуле
36.
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТАЭту реакцию катализирует сложно
организованный пируватдегидрогена
зный комплекс (ПДК). Состоит из трех
типов каталитических протомеров,
образующих три фермента:
Е1 - пируватдекарбоксилазу
(кофермент ТДФ)
Е2 - дигидролипоилтрансацетилазу
(коферменты Липоамид, НS-КоА)
Е3 - дигидролипоилдегидрогена
(коферменты FAD, NAD)
Суммарное уравнение окислительного
декарбоксилирования пирувата
37.
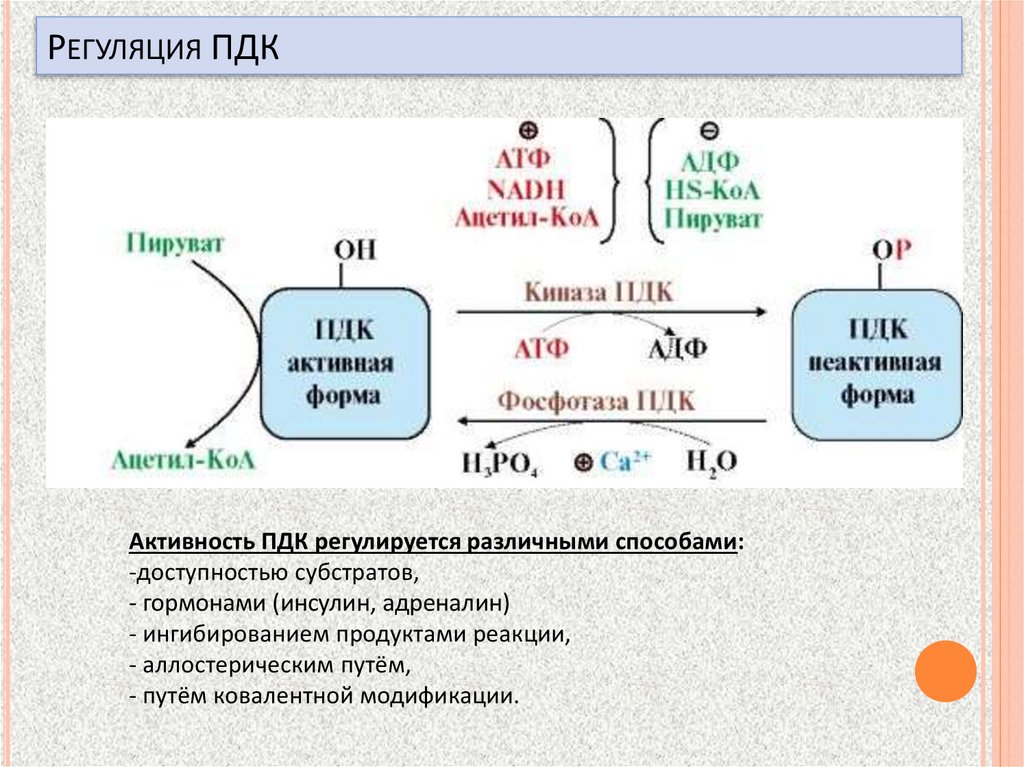
РЕГУЛЯЦИЯ ПДКАктивность ПДК регулируется различными способами:
-доступностью субстратов,
- гормонами (инсулин, адреналин)
- ингибированием продуктами реакции,
- аллостерическим путём,
- путём ковалентной модификации.
38.
Два вспомогательных фермента –киназа и фосфатазаобеспечивают регуляцию активности
пируватдегидрогеназы
киназа активируется при избытке конечного продукта
биологического окисления - АТФ и продуктов ПВКдегидрогеназного комплекса – НАДН и ацетил-S-КоА.
Активная киназа фосфорилирует пируватдегидрогеназу,
инактивируя ее, в результате первая реакция процесса
останавливается.
фосфатаза, активируясь ионами кальция или инсулином,
отщепляет фосфат и активирует пируватдегидрогеназу.
39.
Третий этап – все реакции этапа проходят вмитохондриях.
Ацетил-SКоА включается в реакции цикла трикарбоновых
кислот и окисляется до углекислого газа. Выделенные
атомы водорода восстанавливают НАД и ФАД и после
этого НАДН и ФАДН2 переносят водород в цепь
ферментов дыхательной цепи, расположенную на
внутренней мембране митохондрий. Сюда же отдают
свои атомы водорода молекулы НАДН и ФАДН2,
образованные на втором этапе (гликолиз, окисление
жирных кислот и аминокислот).
В третьем этапе выделяется до 70% всей энергии вещества.
На внутренней мембране митохондрий в результате процесса
под названием "окислительное фосфорилирование"
образуется вода и главный продукт биологического окисления
– АТФ.
40.
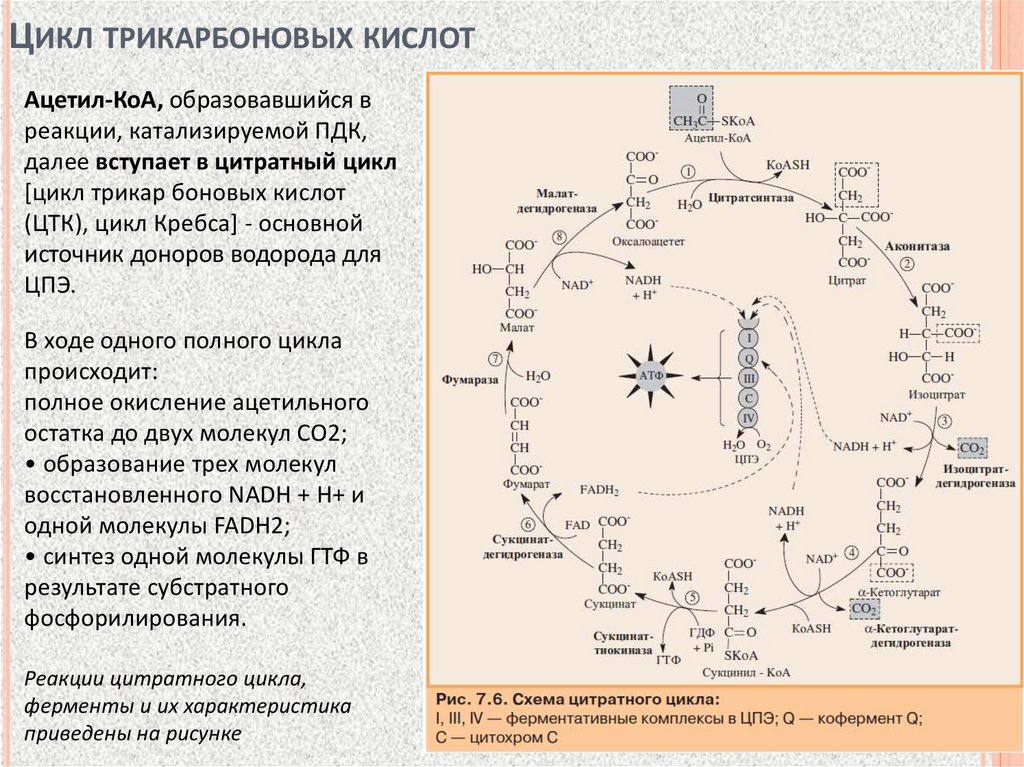
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТАцетил-КоА, образовавшийся в
реакции, катализируемой ПДК,
далее вступает в цитратный цикл
[цикл трикар боновых кислот
(ЦТК), цикл Кребса] - основной
источник доноров водорода для
ЦПЭ.
В ходе одного полного цикла
происходит:
полное окисление ацетильного
остатка до двух молекул СО2;
• образование трех молекул
восстановленного NADH + H+ и
одной молекулы FADH2;
• синтез одной молекулы ГТФ в
результате субстратного
фосфорилирования.
Реакции цитратного цикла,
ферменты и их характеристика
приведены на рисунке
41.
ФУНКЦИИ ЦТК1. Энергетическая (генерация атомов водорода для
работы дыхательной цепи, а именно трех молекул НАДН и
одной молекулы ФАДН2,синтез одной
молекулы ГТФ (эквивалентна АТФ).
2. Анаболическая (в ЦТК образуются предшественник
гема – сукцинил-SКоА, кетокислоты,
способные превращаться в аминокислоты – αкетоглутарат для глутаминовой кислоты, оксалоацетат для
аспарагиновой, лимонная кислота, используемая
для синтеза жирных кислот,оксалоацетат, используемый
для синтеза глюкозы.
42.
Использование метаболитов ОПК в синтезе различныхсоединений:
1, 2, 3 - заменимых аминокислот; 4, 5, 6 - глюкозы; 7 - жирных кислот; 8 гема
43.
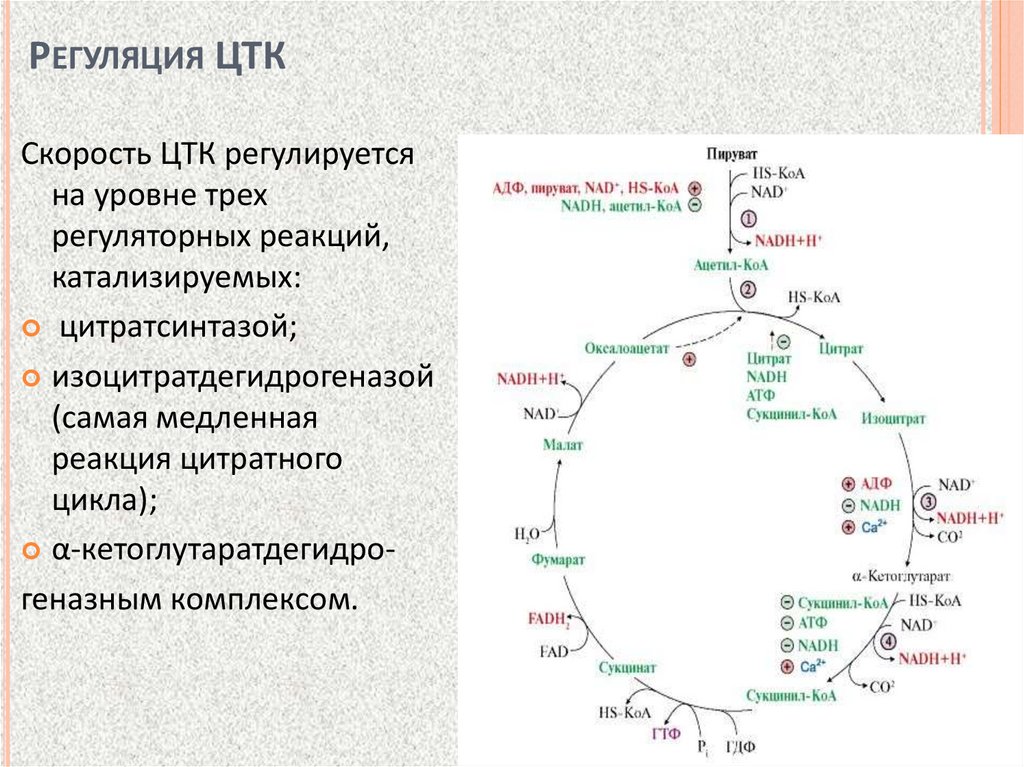
РЕГУЛЯЦИЯ ЦТКСкорость ЦТК регулируется
на уровне трех
регуляторных реакций,
катализируемых:
цитратсинтазой;
изоцитратдегидрогеназой
(самая медленная
реакция цитратного
цикла);
α-кетоглутаратдегидрогеназным комплексом.
44.
ЦИТРАТСИНТАЗАне является аллостерическим ферментом. Активность
фермента регулируется, главным образом, концентрациями
оксалоацетата - субстрата фермента и цитрата - продукта
реакции.
45.
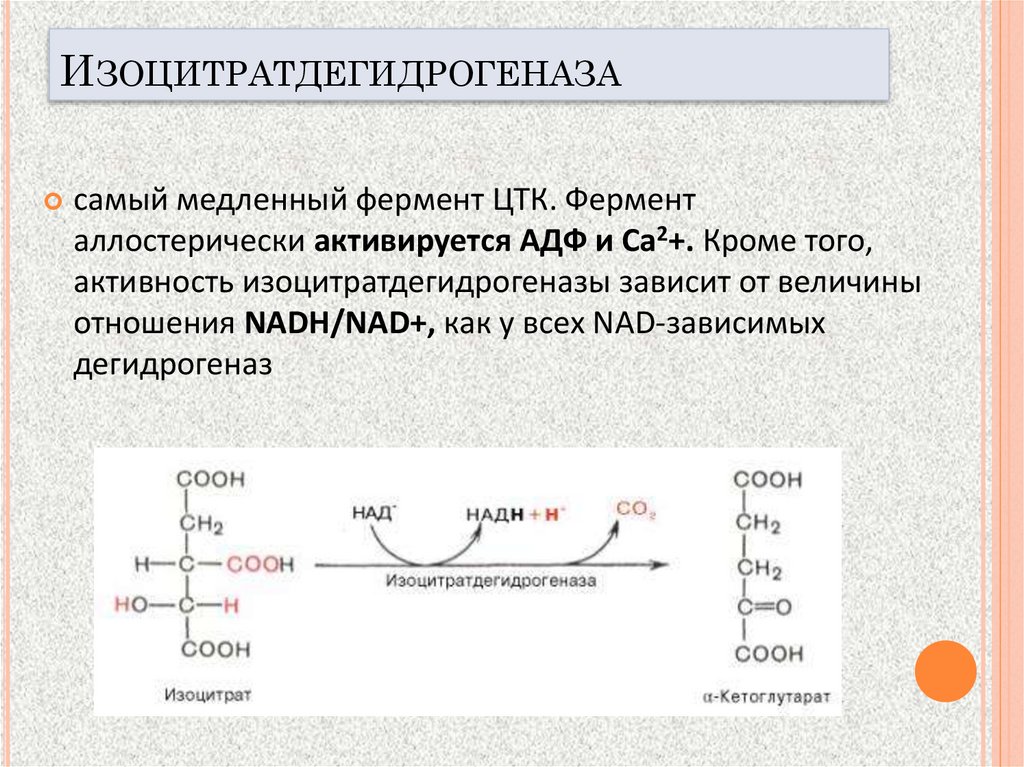
ИЗОЦИТРАТДЕГИДРОГЕНАЗАсамый медленный фермент ЦТК. Фермент
аллостерически активируется АДФ и Са2+. Кроме того,
активность изоцитратдегидрогеназы зависит от величины
отношения NADH/NAD+, как у всех NAD-зависимых
дегидрогеназ
46.
Α-КЕТОГЛУТАРАТДЕГИДРОГЕНАЗНЫЙ КОМПЛЕКСпо структуре и функциям сходен с ПДК. В его состав
входят 3 ферментных комплекса: αкетоглутаратдекарбоксилаза,
дигидролипоилтранссукцинилаза и
дигидролипоилдегидрогеназа.
Набор коферментов аналогичен таковому в ПДК.
Активность фермента зависит от концентраций АТФ и
АДФ, NAD+ и NADH, ингибируется сукцинил-КоА и
активируется Са2+
47.
Суммарное уравнение ЦТК:СН3СО-S-КoA + 2H2O + 3HAД + ФАД+ ГДФ+ Pi →
2CO2 + 3HAДH + ФАДН2+ КoA-SH + ГТФ.
Молекулы НАДН и ФАДН2, образуемые в реакциях окисления
углеводов, жирных кислот, спиртов и аминокислот, далее
поступают в митохондрии, где ферментами дыхательной
цепи осуществляется процесс окислительного
фосфорилирования.
48.
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ– это многоэтапный процесс, происходящий во внутренней
мембране митохондрий и заключающийся в окислении
восстановленных эквивалентов (НАДН и ФАДН2)
ферментами дыхательной цепи (ЦПЭ) и
сопровождающийся синтезом АТФ.
Процесс может происходить только в аэробных условиях.
49.
ПРИНЦИП РАБОТЫ ДЫХАТЕЛЬНОЙ ЦЕПИОбразующиеся в реакциях катаболизма НАДН и
ФАДН2 передают атомы водорода (т.е. протоны водорода и
электроны) на ферменты дыхательной цепи.
Электроны движутся по ферментам дыхательной цепи и
теряют энергию.
Эта энергия используется на выкачивание протонов Н+ из
матрикса в межмембранное пространство.
В конце дыхательной цепи электроны попадают на
кислород и восстанавливают его до воды.
Протоны Н+ стремятся обратно в матрикс и проходят через
АТФ-синтазу.
При этом они теряют энергию, которая используется для
синтеза АТФ.
50.
КОМПЛЕКСЫ ДЫХАТЕЛЬНОЙ ЦЕПИКомплекс I - NADH-дегидрогеназа
Комплекс II - сукцинатдегидрогеназа
Комплекс III - QН2-дегидрогеназа
Комплекс IV - цитохромоксидаза
+ низкомолекулярные переносчики (кофермент Q и
цитохром с)
Комплекс V - фермент АТФ-синтаза
Все компоненты ЦПЭ расположены в митохондриальной мембране в порядке
возрастания редокс-потенциалов; Это обеспечивает последовательное
перемещение электронов от дегидрируемых субстратов на кислород,
сопровождающееся освобождением части свободной энергии электронов
51.
Комплекс I (НАДН-дегидрогеназа) включает ФМН и железосерныйбелок FeS). Комплекс I окисляет НАДН, перенося с него 2 электрона на KoQ.
Комплекс I также перекачивает 4 протона из матрикса в межмембранное
пространство митохондрии.
KoQ (убихинон) - некрупная липофильная молекула. Перемещаясь в
липидном слое мембраны, убихинон обеспечивает передачу электронов
между комплексами I - III и II - III.
Комплекс II (сукцинат-дегидрогеназа) включает ФАД и железосерный
белок. Обеспечивает вход в цепь дополнительных электронов за счёт
окисления сукцината.
Комплекс III (QН2-дегидрогеназа)включает цитохромы b и с1 и
железосерный белок. Комплекс III переносит электроны с убихинона
на цитохром с и перекачивает 2 протона в межмембранное пространство.
Комплекс IV (Цитохром c оксидаза)состоит из цитохромов a и a3, которые,
помимо гема, содержат ионы меди. Комплекс IV катализирует перенос
электронов с молекул цитохрома на O2 и перекачивает 4 протона в
межмембранное пространство.
52.
СОПРЯЖЕНИЕ ТКАНЕВОГО ДЫХАНИЯ И СИНТЕЗА АТФПеренос электронов по ЦПЭ при участии комплексов I, III и IV
сопровождается выделением наибольшего количества энергии. Часть
этой энергии используется для переноса Н+ из матрикса в
межмембранное пространство, в результате чего
возрастает протонный электрохимический потенциал ΔμΗ+
При достижении определенного протонного градиента происходит
активация АТФ-синтазы (комплекс V), в ней открывается канал,
через который протоны возвращаются в матрикс из
межмембранного пространства, а энергия ΔμΗ+ используется для
синтеза АТФ.
Образующаяся АТФ при участии АДФ-АТФ-транслоказы транспортируется из
матрикса на наружную сторону мембраны и попадает в цитозоль. Одновременно
та же транслоказа переносит АДФ в обратном направлении, из цитозоля в
матрикс митохондрии.
53.
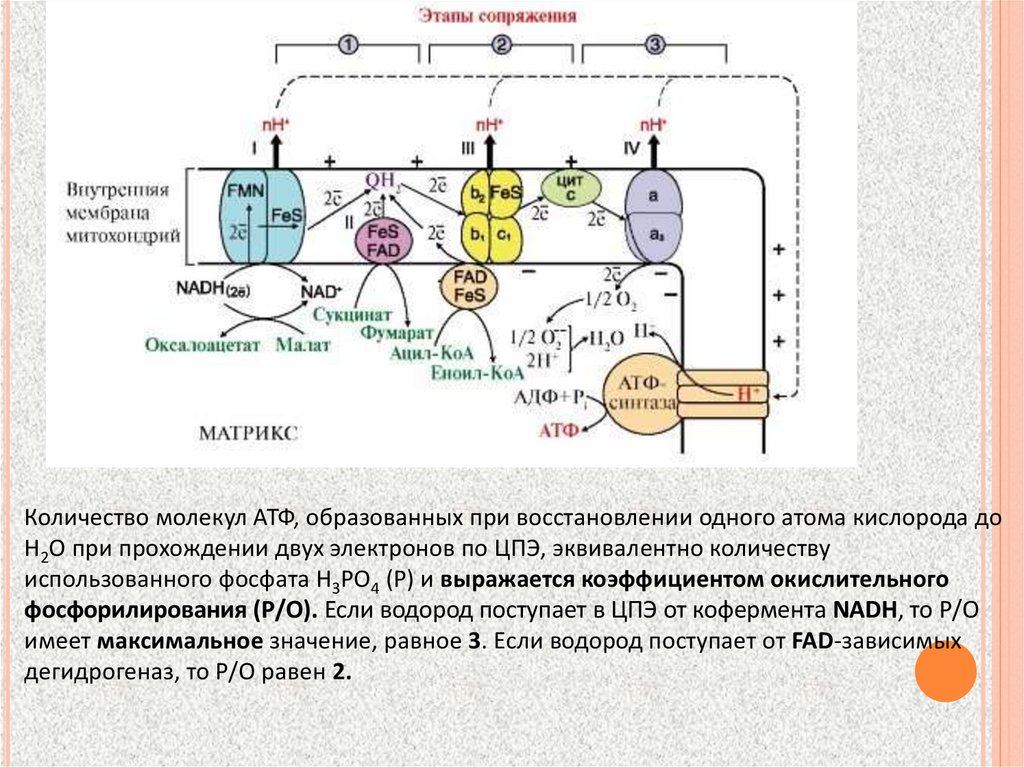
Количество молекул АТФ, образованных при восстановлении одного атома кислорода доН2О при прохождении двух электронов по ЦПЭ, эквивалентно количеству
использованного фосфата Н3РО4 (Р) и выражается коэффициентом окислительного
фосфорилирования (Р/О). Если водород поступает в ЦПЭ от кофермента NADH, то Р/О
имеет максимальное значение, равное 3. Если водород поступает от FAD-зависимых
дегидрогеназ, то Р/О равен 2.
54.
ДЫХАТЕЛЬНЫЙ КОНТРОЛЬУскорение тканевого дыхания и окислительного
фосфорилирования при повышении концентрации
AДФ называется дыхательным контролем.
Гипоэнергетические состояния подразделяются на:
- алиментарные (голодание, авитаминоз);
- гипоксические:
- нарушение поступления кислорода в кровь (легочная гипоксия),
- нарушение транспорта кислорода в ткани (гемодинамическая
(кровопотеря, шок, пороки сердца) и гемоглобиновая (патология
гемоглобина, блокирование его ядами) гипоксия),
- митохондриальные (затруднение использования кислорода в клетках) нарушение функций митохондрий ингибиторами ферментов дыхательной
цепи, разобщителями окисления и фосфорилирования,
мембранотропными веществами.
55.
РАЗОБЩЕНИЕ ОКИСЛЕНИЯ И ФОСФОРИЛИРОВАНИЯРазобщители - липофильные вещества, которые способны
принимать протоны и переносить их через внутреннюю мембрану
митохондрий, минуя его протонный канал. В результате этого
снижается протонный градиент и прекращается синтез АТФ.
Разобщители (протонофоры) бывают:
- естественные – продукты перекисного окисления липидов,
жирных кислот с длинной цепью; большие дозы тиреоидных
гормонов;
- искусственные - динитрофенол, эфир, анестетики, антибиотики
(грамицидин, валиномицин)
При действии разобщающих факторов большая часть энергии
выделяется в виде теплоты, количество АДФ и поглощение
кислорода при этом увеличиваются. Следствием эффекта
протонофоров является возрастание катаболизма жиров и
углеводов в клетке.
56.
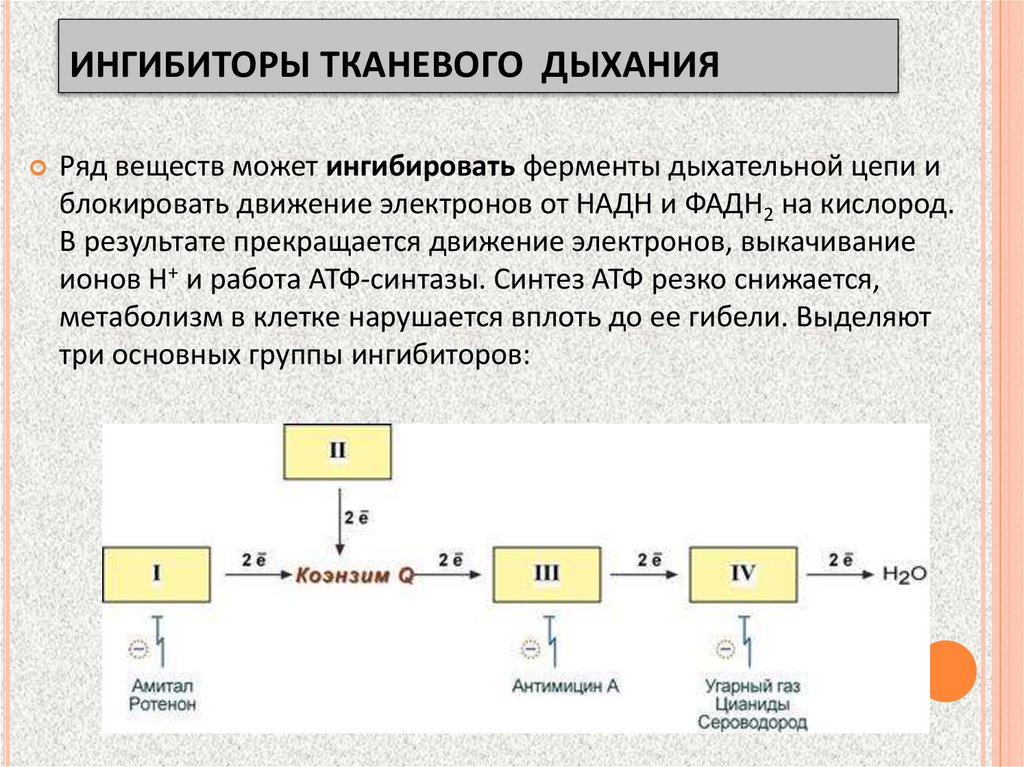
ИНГИБИТОРЫ ТКАНЕВОГО ДЫХАНИЯРяд веществ может ингибировать ферменты дыхательной цепи и
блокировать движение электронов от НАДН и ФАДН2 на кислород.
В результате прекращается движение электронов, выкачивание
ионов Н+ и работа АТФ-синтазы. Синтез АТФ резко снижается,
метаболизм в клетке нарушается вплоть до ее гибели. Выделяют
три основных группы ингибиторов:
57.
Ответьте на вопрос:Амитал, ротенон, угарный газ,
цианиды – ингибиторы
дыхательной цепи
Почему барбитураты (амитал
натрия и др.) используются
в медицинской практике
как снотворные средства, а
цианид калия является
смертельным ядом?
58.
ЗАКЛЮЧЕНИЕЖивые организмы осуществляют непрерывную и
постоянную связь с окружающей средой в процессе
метаболизма. Эндокринная система обеспечивает
регуляцию и интеграцию метаболизма в разных
тканях в ответ на изменения условий внешней и
внутренней среды.
59.
ЛИТЕРАТУРАБиологическая химия с упражнениями и задачами:
учебник для студентов ВУЗов / ред. С. Е. Северин. М.: ГЭОТАР-Медиа, 2010. - 380 с.
Березов Т.Т. Биологическая химия: учебник для студ.
мед. ВУЗов [Рекомендовано УМО] / Т. Т. Березов, Б.
Ф. Коровкин. -3-е изд., перераб. и доп. -М.:
Медицина, 2004. -704 с.
Биохимия для студента (Электронный
ресурс):сайт. – Режим доступа:
https://biokhimija.ru/- Свободный доступ