





































 biology
biologySimilar presentations:










Мышечная ткань
1.
МИНИСТЕРСТВОНАУКИ И ВЫСШЕГО ОБРАЗОВАНИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ ТКАНЬ
МЕЗЕНХИМА
—
ЭМБРИОНАЛЬНАЯ
СОЕДИНИТЕЛЬНАЯ
ФГАОУ ВО «КРЫМСКИЙ ФЕДЕРАЛЬНЫЙ УНИВЕРСИТЕТ ИМЕНИ В.И.ВЕРНАДСКОГО»
Институт «МЕДИЦИНСКАЯ АКАДЕМИЯ ИМЕНИ С.И. ГЕОРГИЕВСКОГО»
1
Кафедра гистологии и эмбриологии
Лекцию подготовил доцент Лугин И.А.
2.
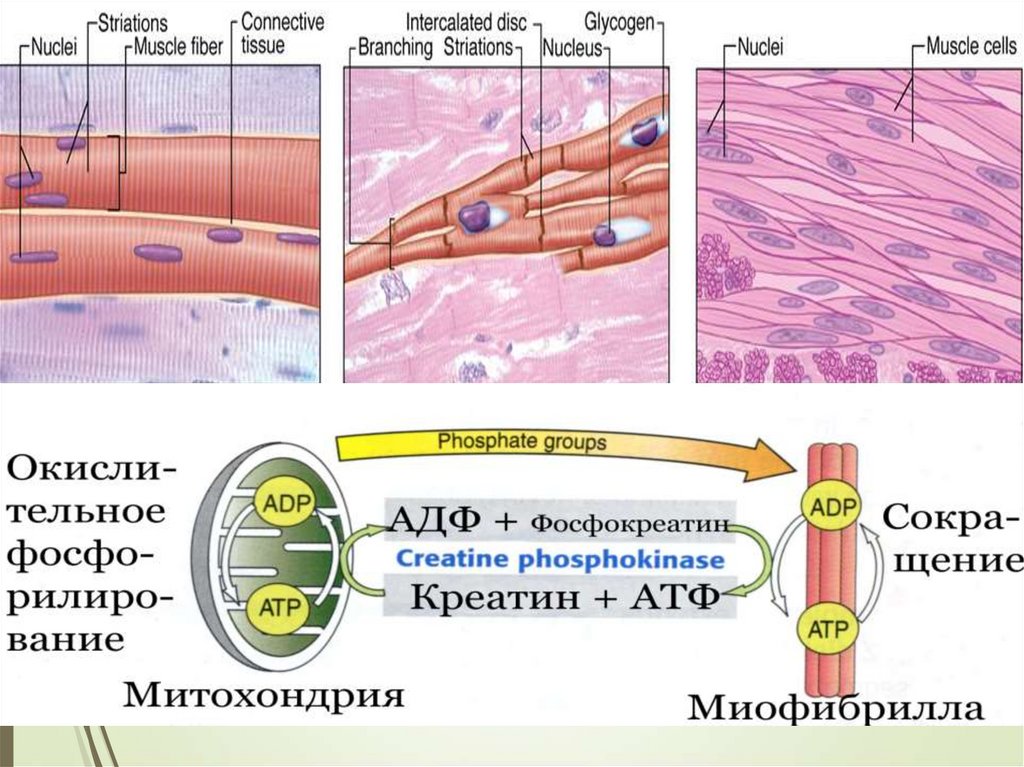
МЫШЕЧНЫЕ ТКАНИ2
Мышечными тканями называют группу
тканей, различных по строению и
происхождению, но сходных по
способности к выраженным
сокращениям
Сократимость – главный
функциональный признак мышечных
тканей, благодаря которому мышечные
ткани могут перемещать тело человека
или его части в пространстве – что
является главной функцией!
Для всех типов мышечных тканей
характерна удлиненная форма клеток
или структурных компонентов, а
способность к сокращению реализуется
на основе актино-миозиновых аппаратов
различной степени сложности –
специализированных органелл миофибрилл
3.
3СОКРАТИТЕЛЬНЫЙ АППАРАТ
Специальные сократительные органеллы —
миофиламенты обеспечивают сокращение,
которое возникает при взаимодействии в них
двух основных фибриллярных белков —
актина и миозина при обязательном участии
ионов кальция (Са2+)
Митохондрии обеспечивают эти процессы
энергией.
Запас источников энергии образуют гликоген
и липиды.
Миоглобин — это белок-пигмент (наподобие
гемоглобина), обеспечивающий связывание
кислорода и создание его запаса на момент
сокращения мышцы, когда сдавливаются
кровеносные сосуды (и поступление
кислорода при этом резко падает).
4.
5.
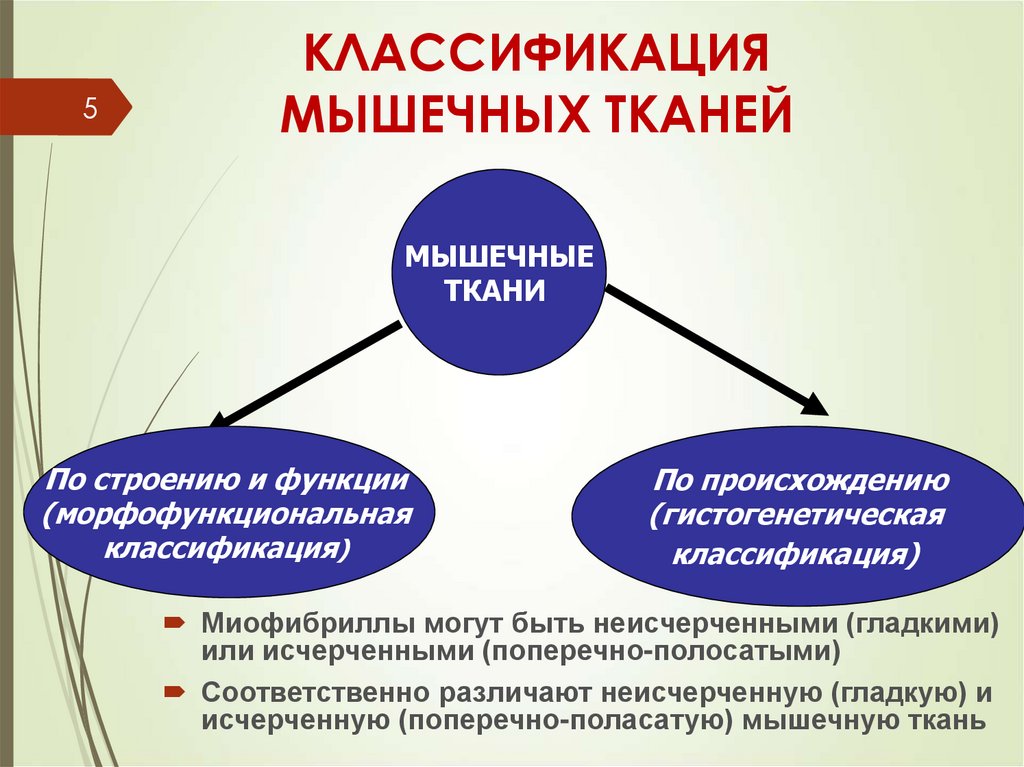
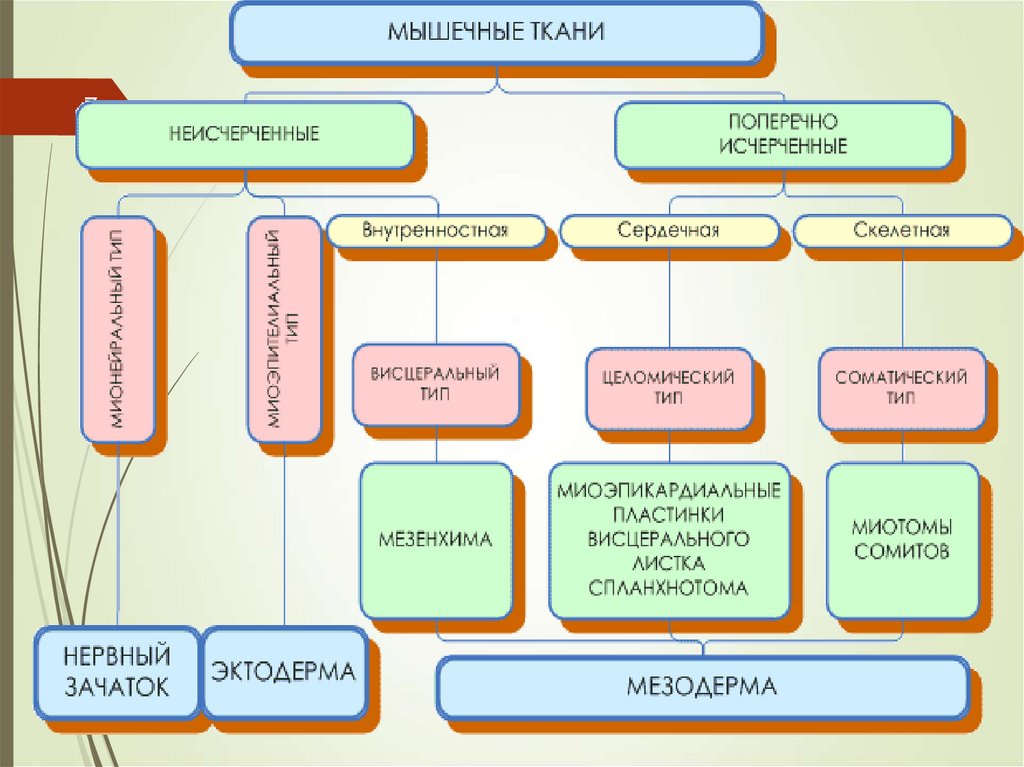
5КЛАССИФИКАЦИЯ
МЫШЕЧНЫХ ТКАНЕЙ
МЫШЕЧНЫЕ
ТКАНИ
По строению и функции
(морфофункциональная
классификация)
По происхождению
(гистогенетическая
классификация)
Миофибриллы могут быть неисчерченными (гладкими)
или исчерченными (поперечно-полосатыми)
Соответственно различают неисчерченную (гладкую) и
исчерченную (поперечно-поласатую) мышечную ткань
6.
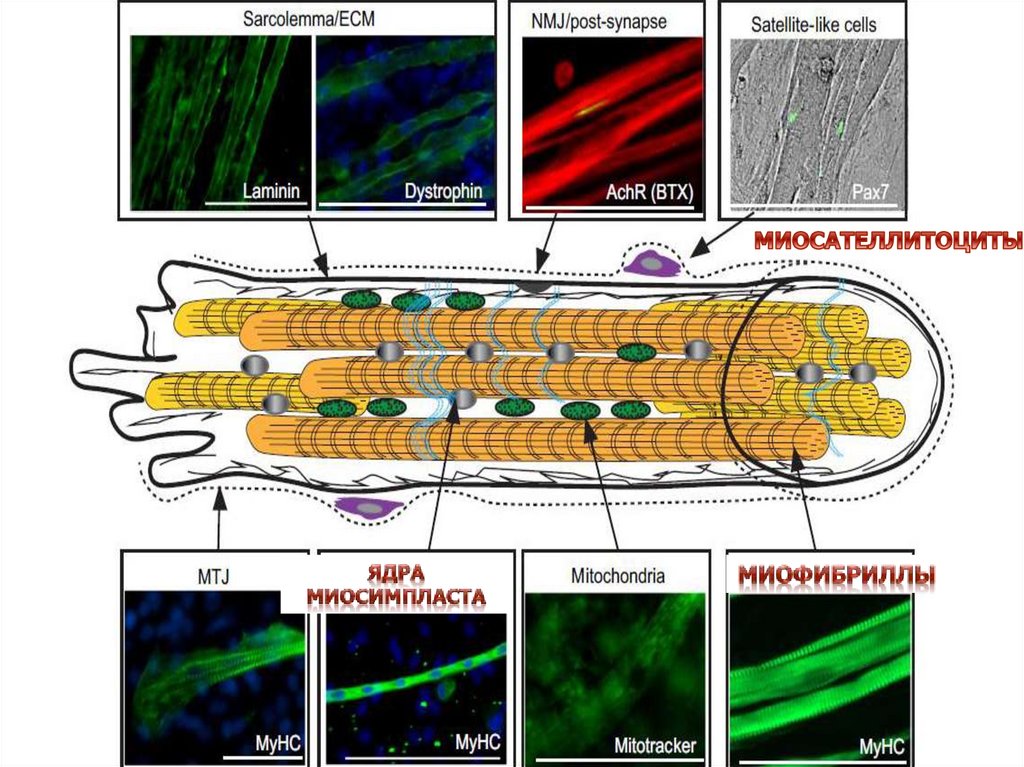
Основным источникомфизиологической и
репаративной регенерации
скелетной мышечной ткани
являются миосателлитоциты,
представляющие собой
перспективный материал для
разработки методов клеточной
терапии ряда мышечных
заболеваний.
Следует отметить, что в 30–40 гг.
прошлого века отечественный
гистолог- эволюционист Н.Г.
Хлопин на основе собственных
научных наблюдений
высказывал предположение о
функциональном различии ядер
в составе мышечных волокон
Онто-филлогенетическая (МВ)
классификация 1947г.
7.
78.
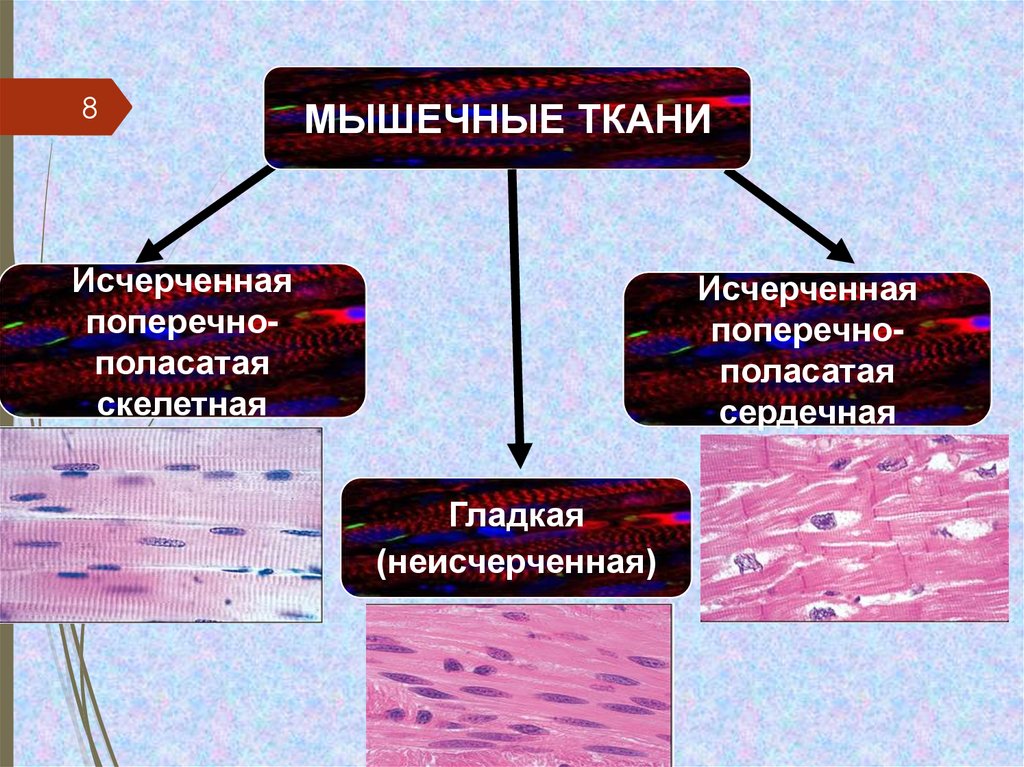
8МЫШЕЧНЫЕ ТКАНИ
Исчерченная
поперечнополасатая
скелетная
Исчерченная
поперечнополасатая
сердечная
Гладкая
(неисчерченная)
9.
910.
10ОСОБЕННОСТИ МЫШЕЧНЫХ ТКАНЕЙ
Удлиненная форма структурных элементов
Миофилламенты – располагаются продольно
(продольная исчерченность
Сократительный аппарат связан с
цитоскелетом и плазмолеммой
Мышечные сокращения требуют много АТФ и
ионов Са2+
Много трофических включений(гликоген и
липиды)
Присутствует МИОГЛОБИН активирующий
окислительное фосфорилирование
Развиты органеллы специального назначения
Саркоплазматическая сеть, кавеолы,
миофибриллы
11.
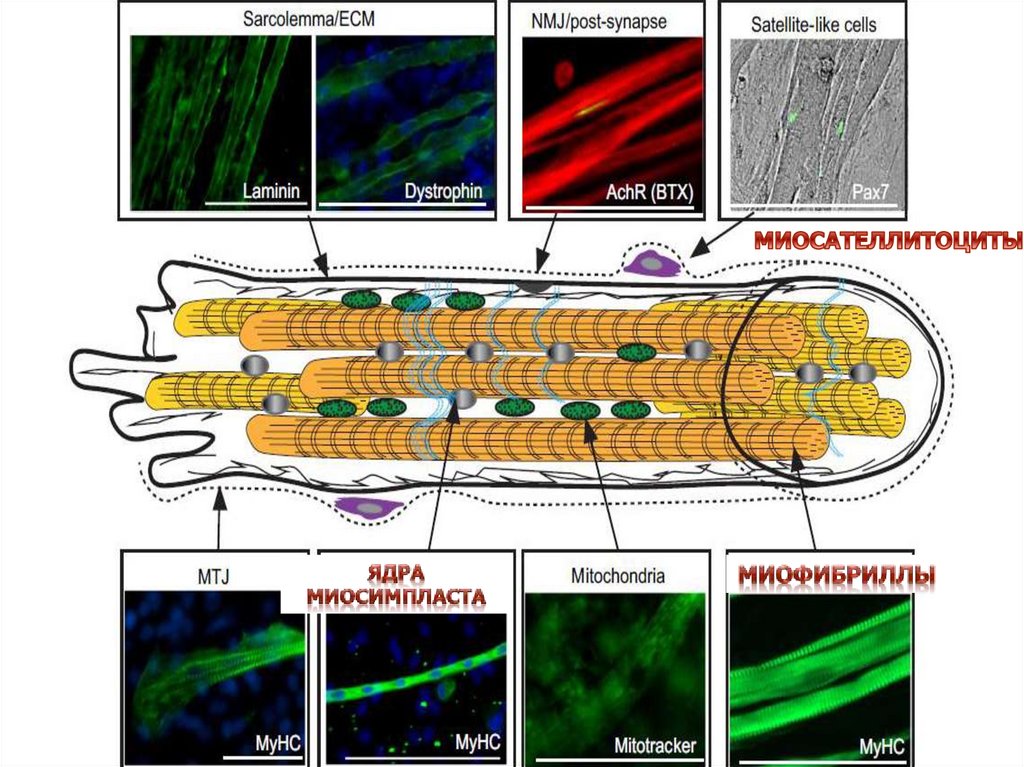
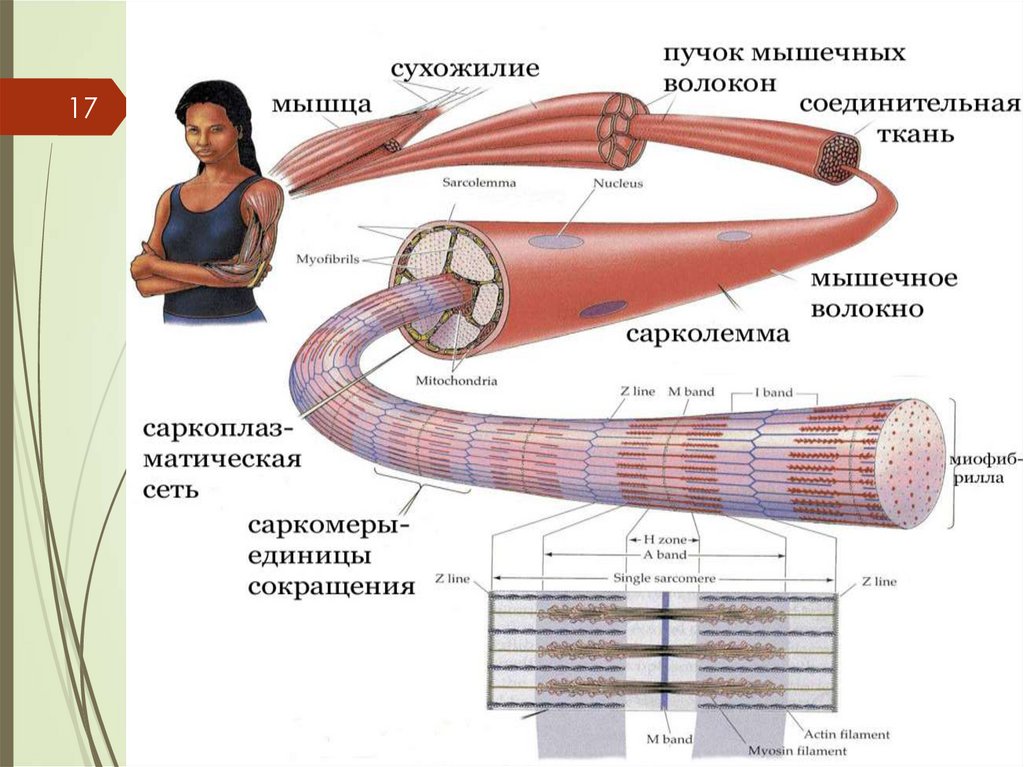
СТРОЕНИЕ МЫШЦ11
СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ
ЕДИНИЦЕЙ СКЕЛЕТНОЙ МЫШЕЧНОЙ
ТКАНИ – МЫШЕЧНОЕ ВОЛОКНО
СОСТОИТ ИЗ МИОСИМПЛАСТА И
МИОСАТЛЛИТОЦИТОВ ПОКРЫТЫХ
ОБЩЕЙ БАЗАЛЬНОЙ МЕМБРАНОЙ
ДЛИНА САНТИМЕТРЫ,
ШИРИНА -50-100 мкм
12.
13.
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬКомплекс, состоящий из плазмолеммы
миосимпласта и базальной мембраны,
называют сарколеммой
Миосимпласт имеет множество
продолговатых ядер, расположенных
непосредственно под сарколеммой.
(количество в одном симпласте может
достигать нескольких десятков тысяч)
У полюсов ядер располагаются
органеллы общего значения — аппарат
Гольджи и небольшие фрагменты
гранулярной эндоплазматической сети
Миофибриллы заполняют основную часть
миосимпласта и расположены продольно
14.
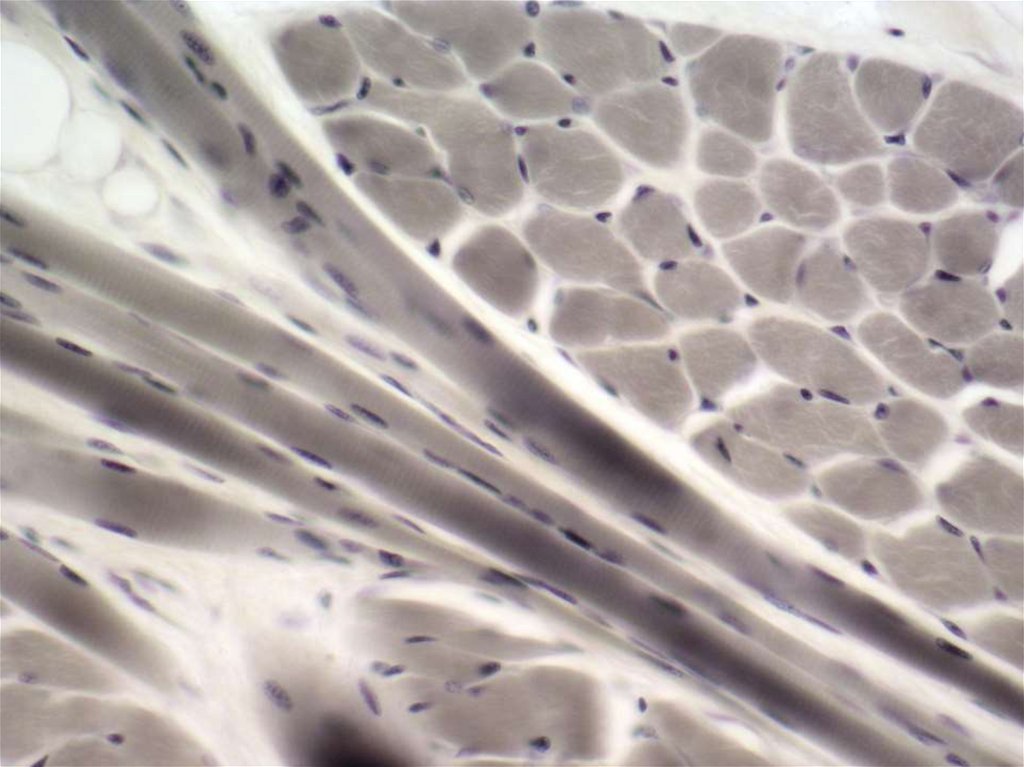
14ПРОДОЛЬНЫЙ СРЕЗ ПОПЕРЕЧНО-ПОЛОСАТОЙ
МЫШЕЧНОЙ ТКАНИ. ОКРАСКА : пикросириус
+поляризованный свет. В ЛЕВОЙ ЧАСТИ ВИДНО
ПРОНИКНОВЕНИЕ КОЛЛАГЕНОВЫХ ВОЛОКОН В
МЫШЦУ.
15.
1516.
17.
1718.
1819.
19МЫШЕЧНЫЕ
ТКАНИ
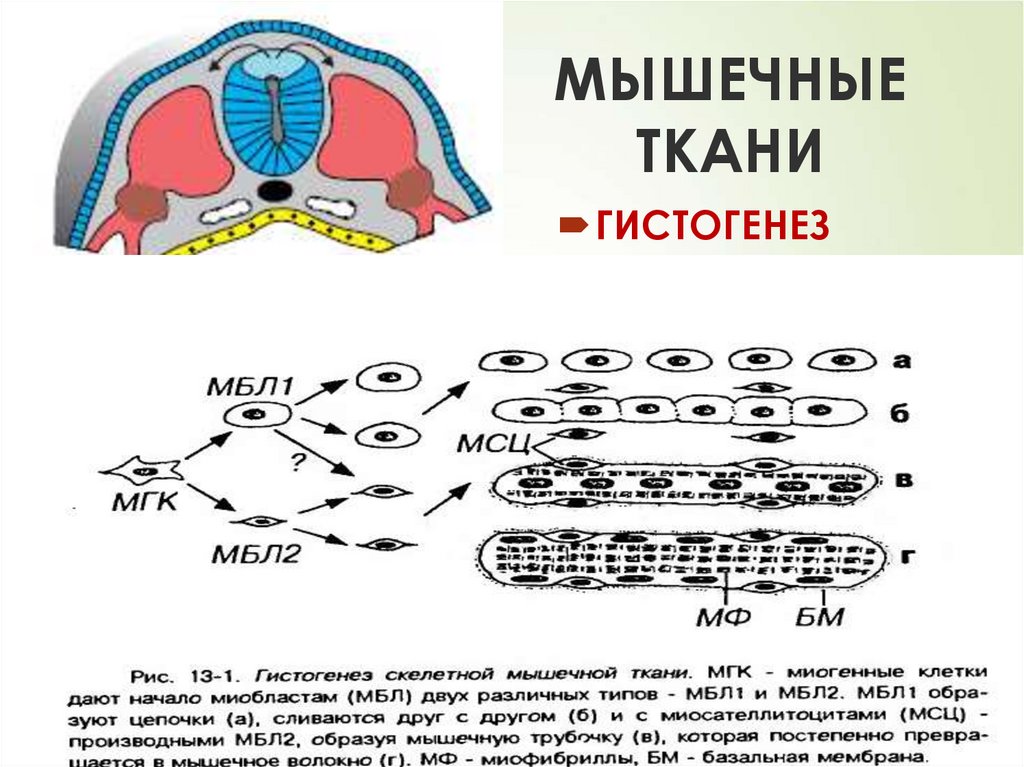
ГИСТОГЕНЕЗ
20.
ИСТОЧНИКРАЗВИТИЯ
20
СТВОЛОВЫЕ КЛЕТКИ
МИОТОМОВ
ПРОМИОБЛАСТЫ
ФОРМИРОВАНИЕ
АУТОХТОННЫХ МЫШЦ
ДРУГИЕ МИГРИРУЮТ В
МЕЗЕНХИМУ
ФОРМИРУЮТ МЫШЦЫ
ТЕЛА
ДВЕ КЛЕТОЧНЫЕ ЛИНИИ :
МИОСИМПЛАСТОВ
И МИОСАТЕЛЛИТОЦИТОВ
21.
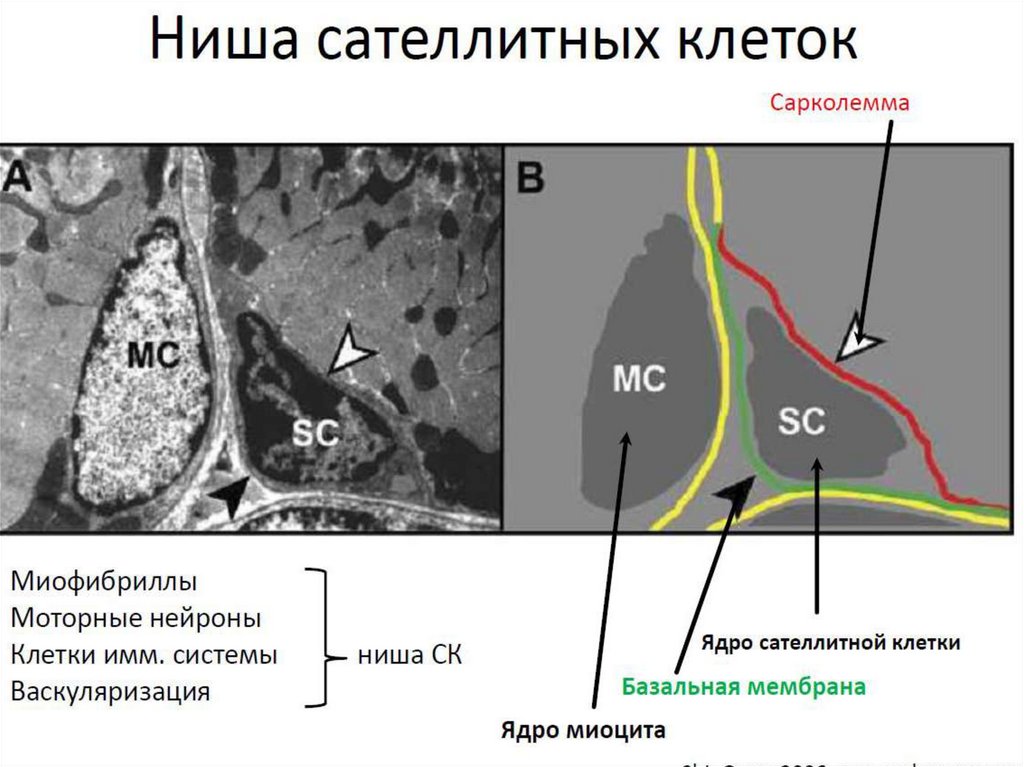
Расположение миосателлитоцитов подбазальной мембраной мышечного волокна
играет важную роль в поддержании их
пролиферативного покоя
По длине мышечного волокна
миосателлитоциты распределены
неравномерно:
обнаружены места наибольшей
концентрации этих клеток
Отмечается частое расположение
миосателлитоцитов вблизи нервно-мышечных
синапсов, мышечно-сухожильных
соединений, кровеносных капилляров
Таким образом это участки стволовой ниши
22.
23.
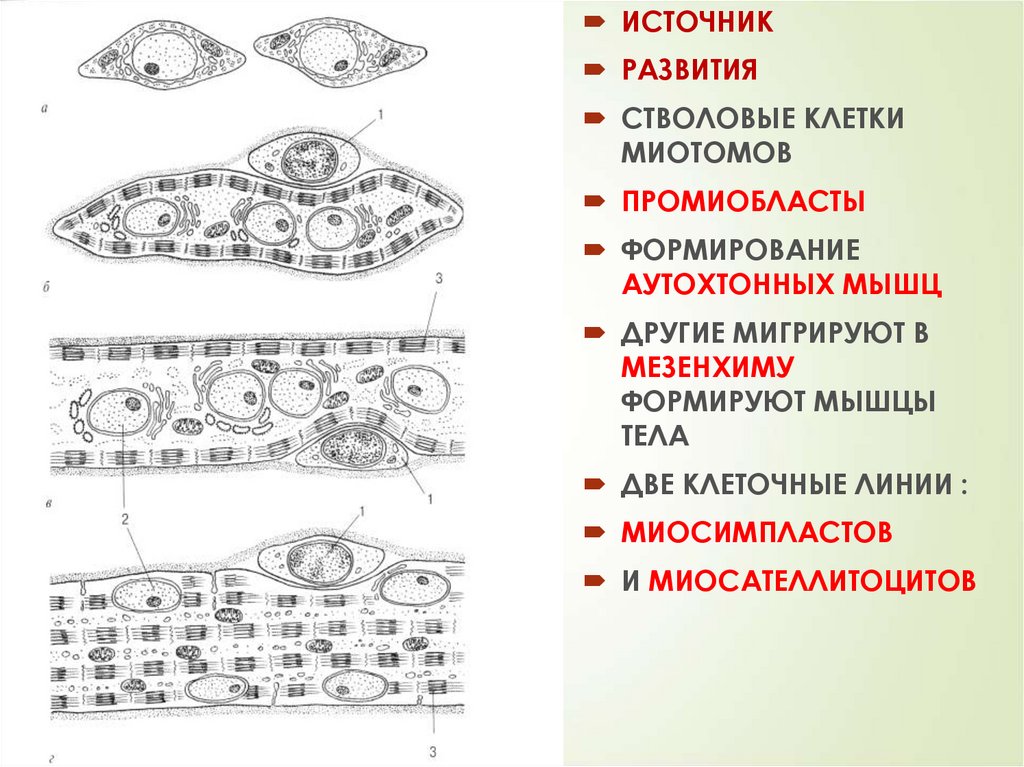
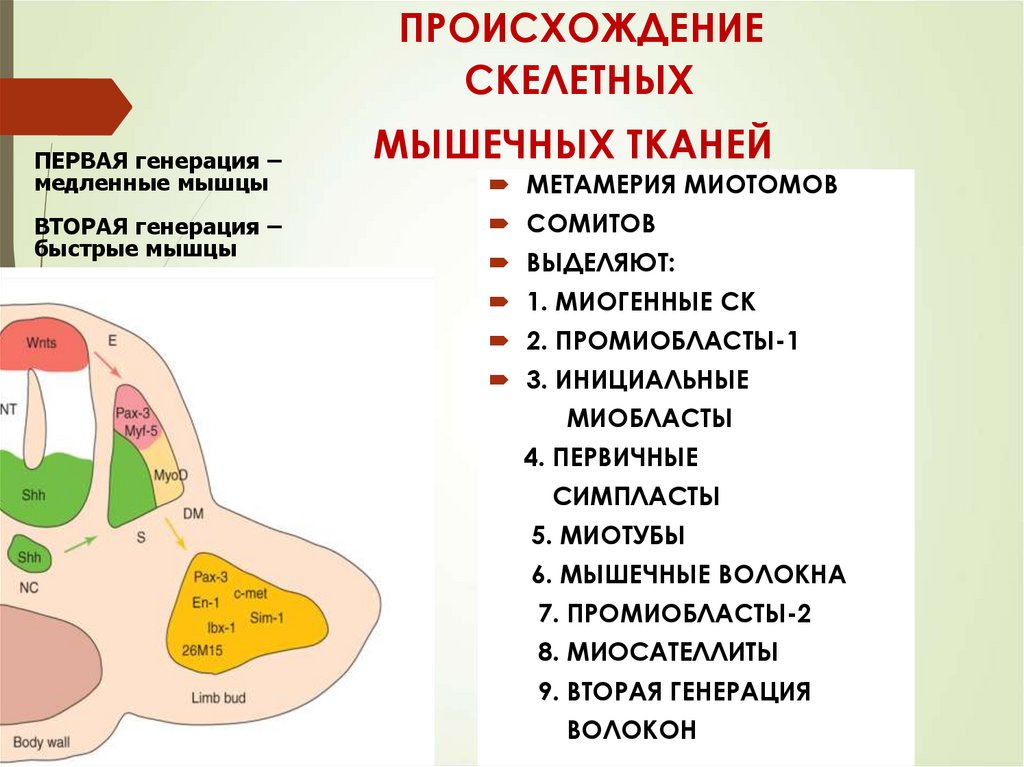
ПРОИСХОЖДЕНИЕСКЕЛЕТНЫХ
ПЕРВАЯ генерация –
медленные мышцы
ВТОРАЯ генерация –
быстрые мышцы
МЫШЕЧНЫХ ТКАНЕЙ
МЕТАМЕРИЯ МИОТОМОВ
СОМИТОВ
ВЫДЕЛЯЮТ:
1. МИОГЕННЫЕ СК
2. ПРОМИОБЛАСТЫ-1
3. ИНИЦИАЛЬНЫЕ
МИОБЛАСТЫ
4. ПЕРВИЧНЫЕ
СИМПЛАСТЫ
5. МИОТУБЫ
6. МЫШЕЧНЫЕ ВОЛОКНА
7. ПРОМИОБЛАСТЫ-2
8. МИОСАТЕЛЛИТЫ
9. ВТОРАЯ ГЕНЕРАЦИЯ
ВОЛОКОН
24.
АППАРАТЫ МЫШЕЧНОГО ВОЛОКНА24
а)Трофический аппарат - совокупность
ядерно - плазматических структур.
б)Сократительный аппарат, представленный
поперечнополосатыми миофибриллами.
в)Опорный аппарат мышечного волокна
включает особые элементы цитоскелета,
обеспечивающие высокоупорядоченное
расположение миофиламентов и
миофибрилл внутри волокна, а также
связанную с ними сарколемму соединяющую
мышечное волокно с сухожилием, на которое
передается усилие, развиваемое волокном
при сокращении.
г) Аппарат передачи возбуждения
(САРКОПЛАЗМИТИЧЕСКИЙ)
г) Энергетический – митохондрии
д) Лизосомальный – регенераторный
25.
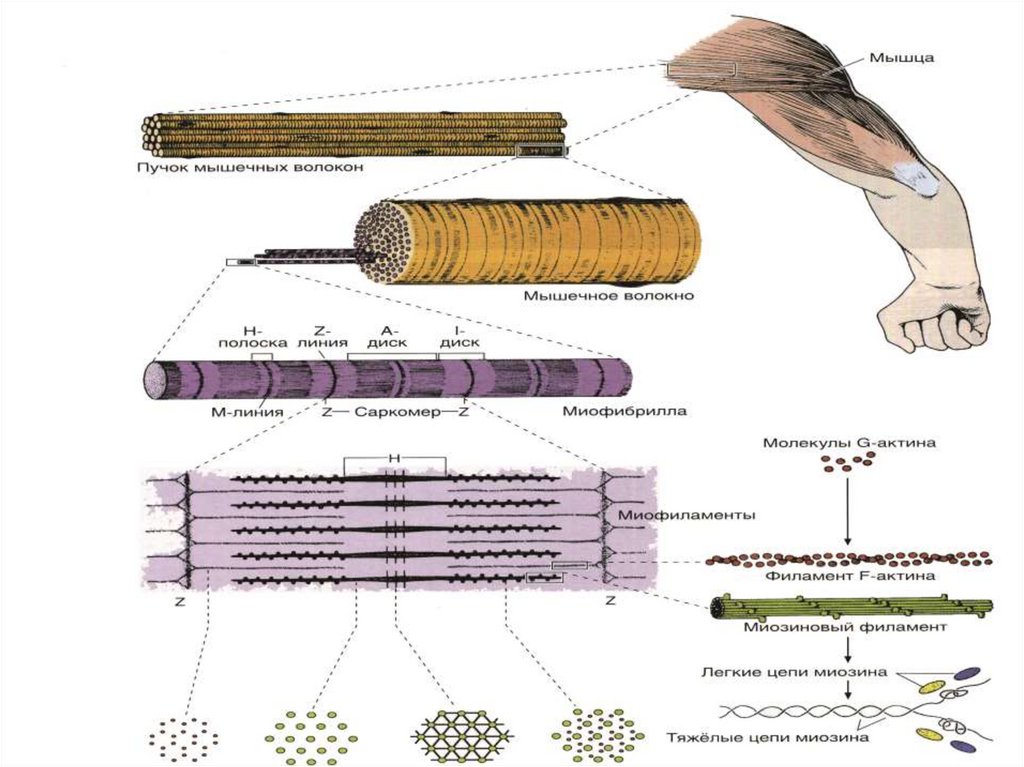
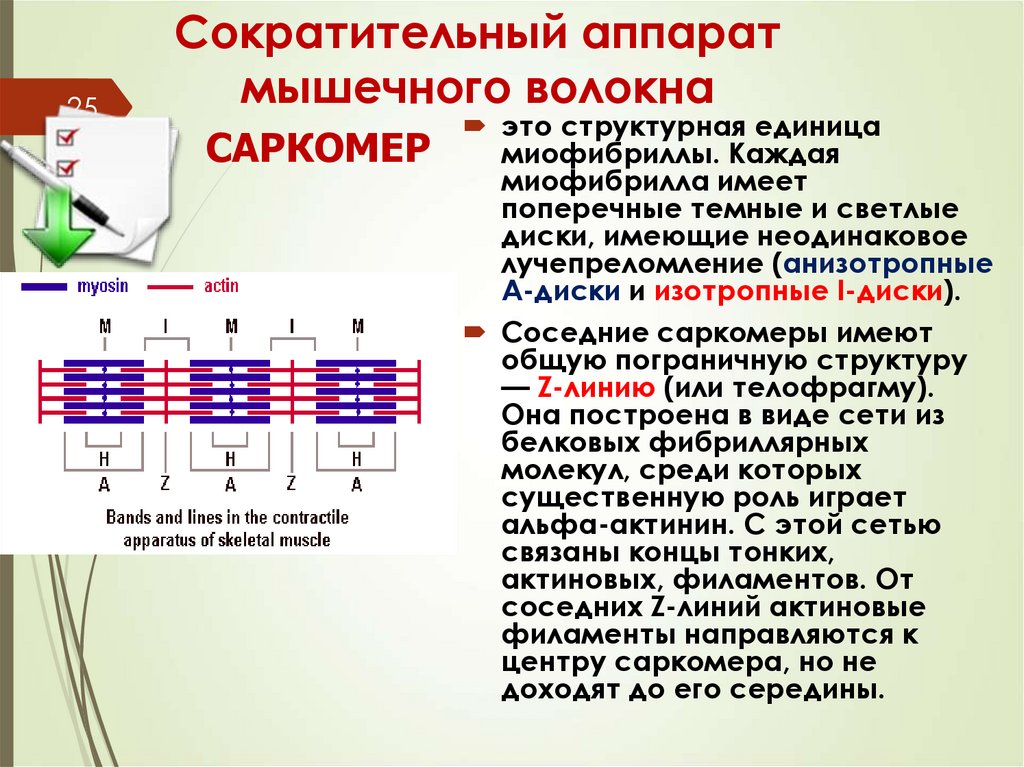
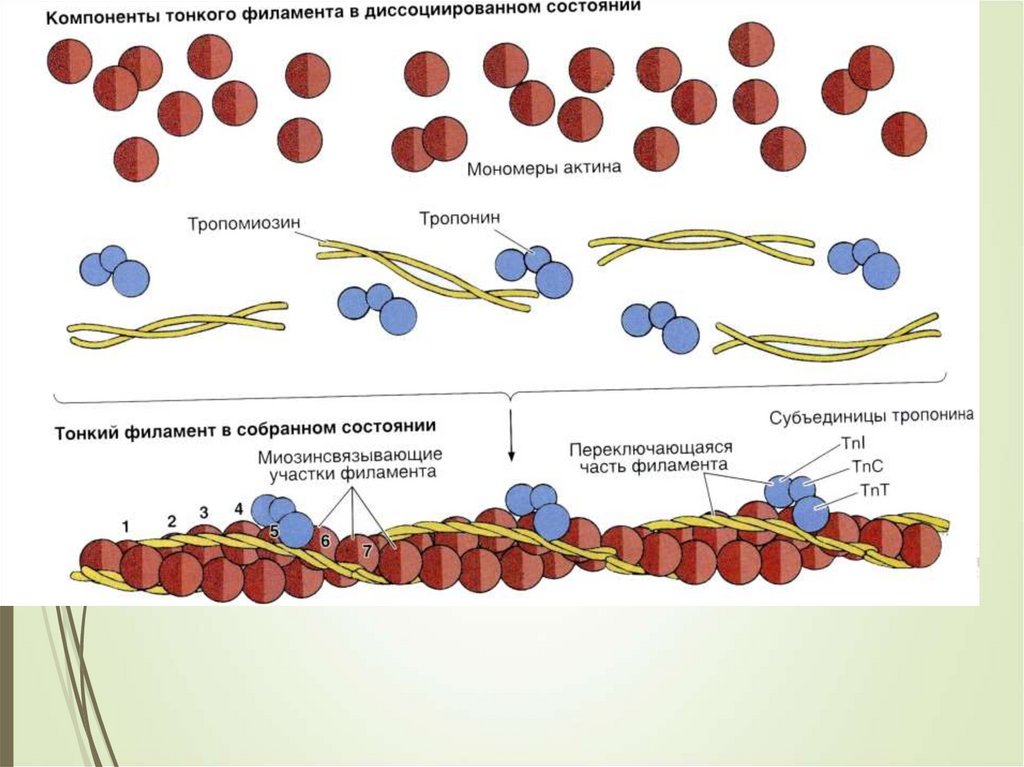
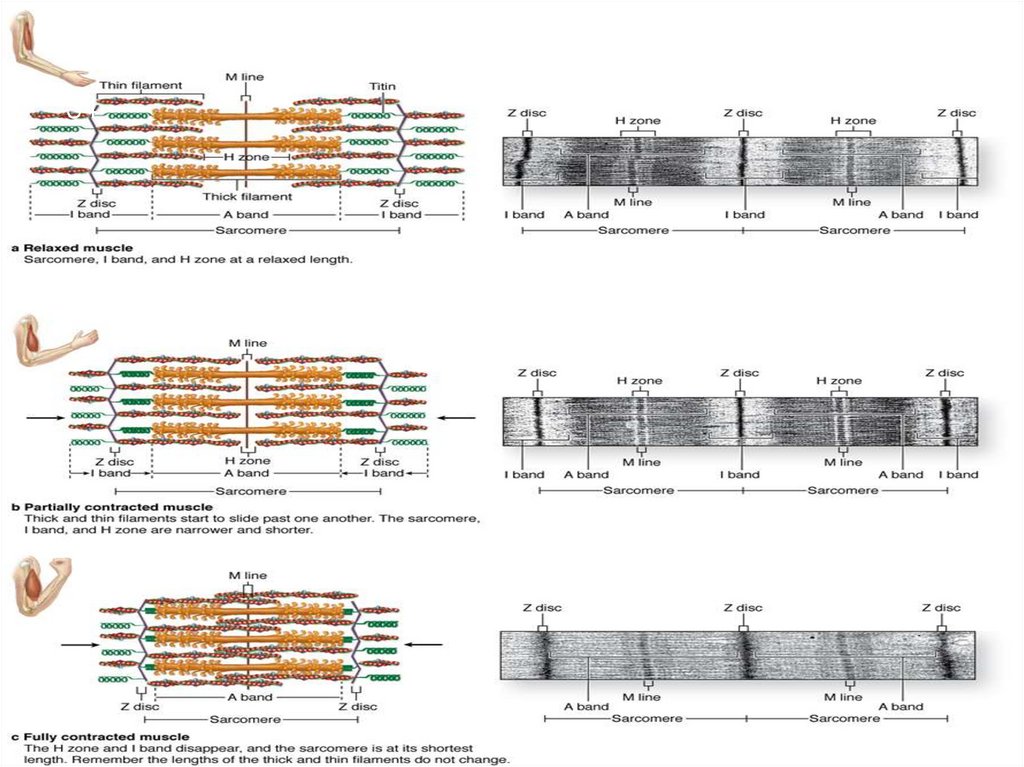
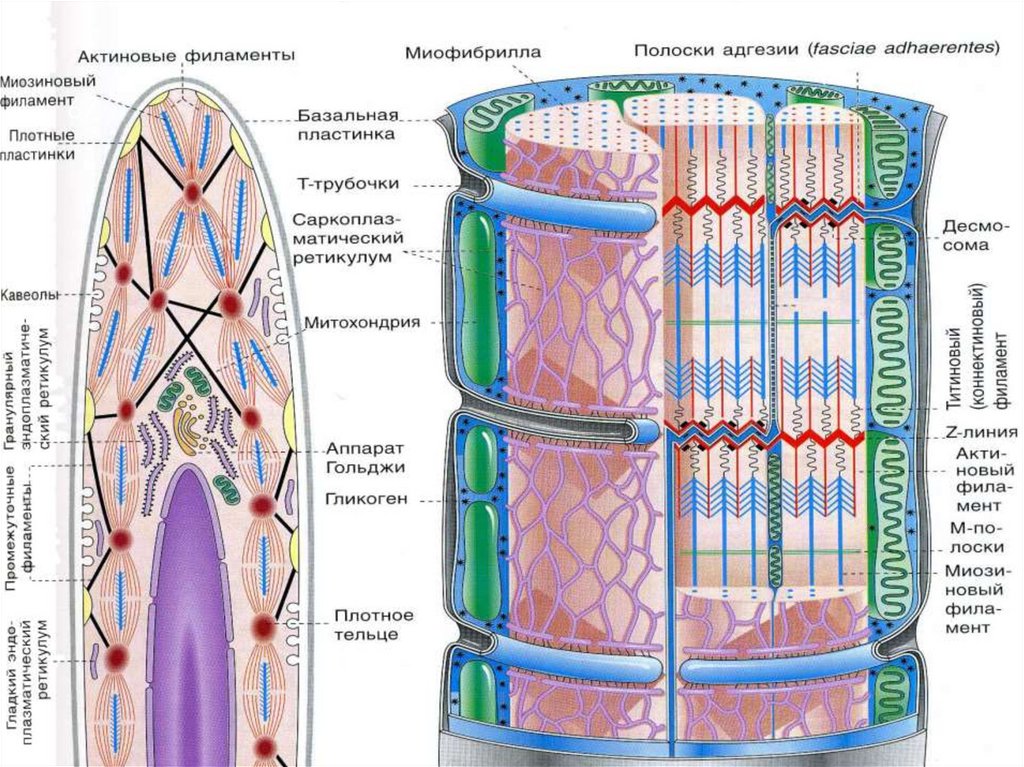
25Сократительный аппарат
мышечного волокна
САРКОМЕР
это структурная единица
миофибриллы. Каждая
миофибрилла имеет
поперечные темные и светлые
диски, имеющие неодинаковое
лучепреломление (анизотропные
A-диски и изотропные I-диски).
Соседние саркомеры имеют
общую пограничную структуру
— Z-линию (или телофрагму).
Она построена в виде сети из
белковых фибриллярных
молекул, среди которых
существенную роль играет
альфа-актинин. С этой сетью
связаны концы тонких,
актиновых, филаментов. От
соседних Z-линий актиновые
филаменты направляются к
центру саркомера, но не
доходят до его середины.
26.
2627.
2728.
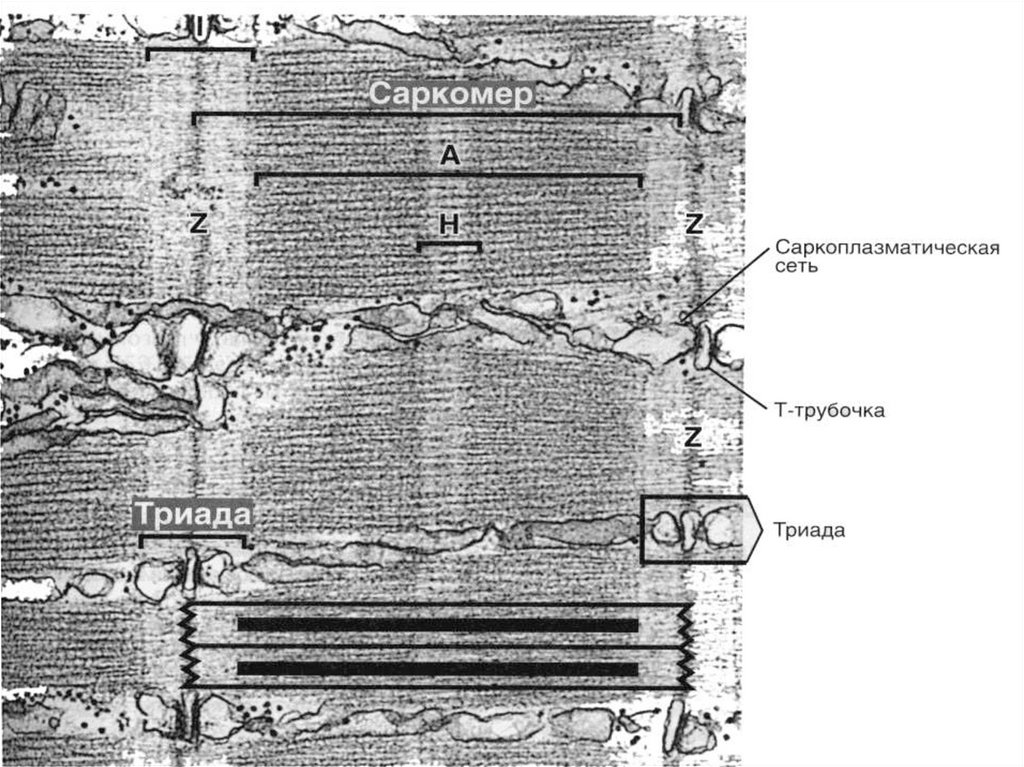
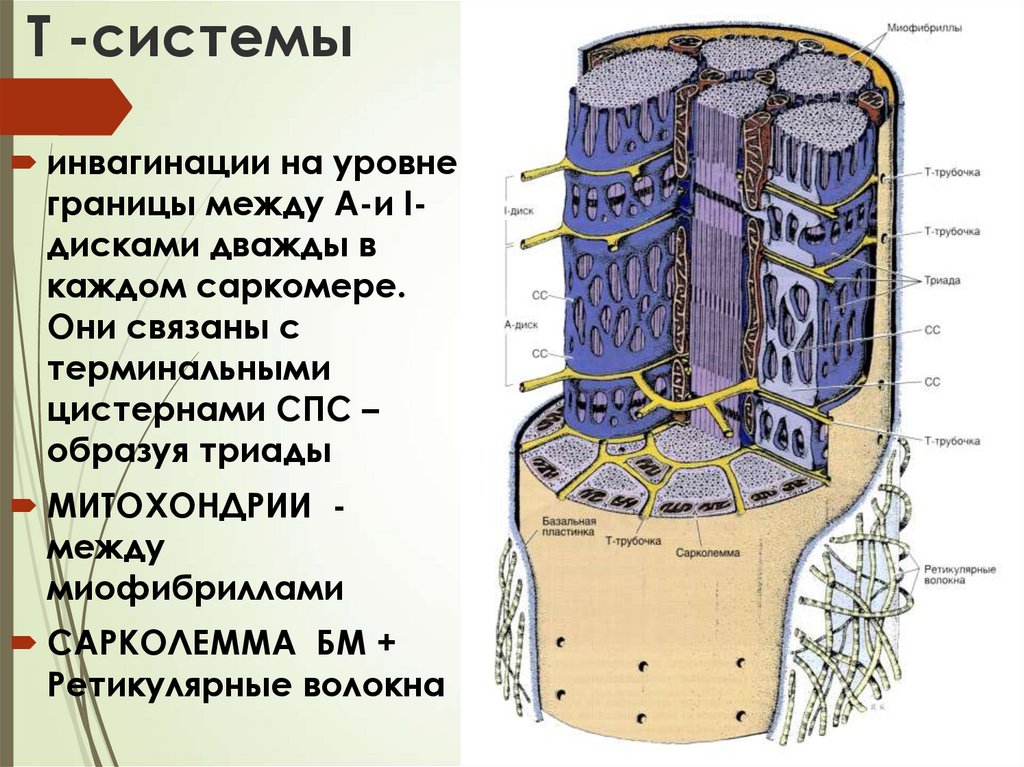
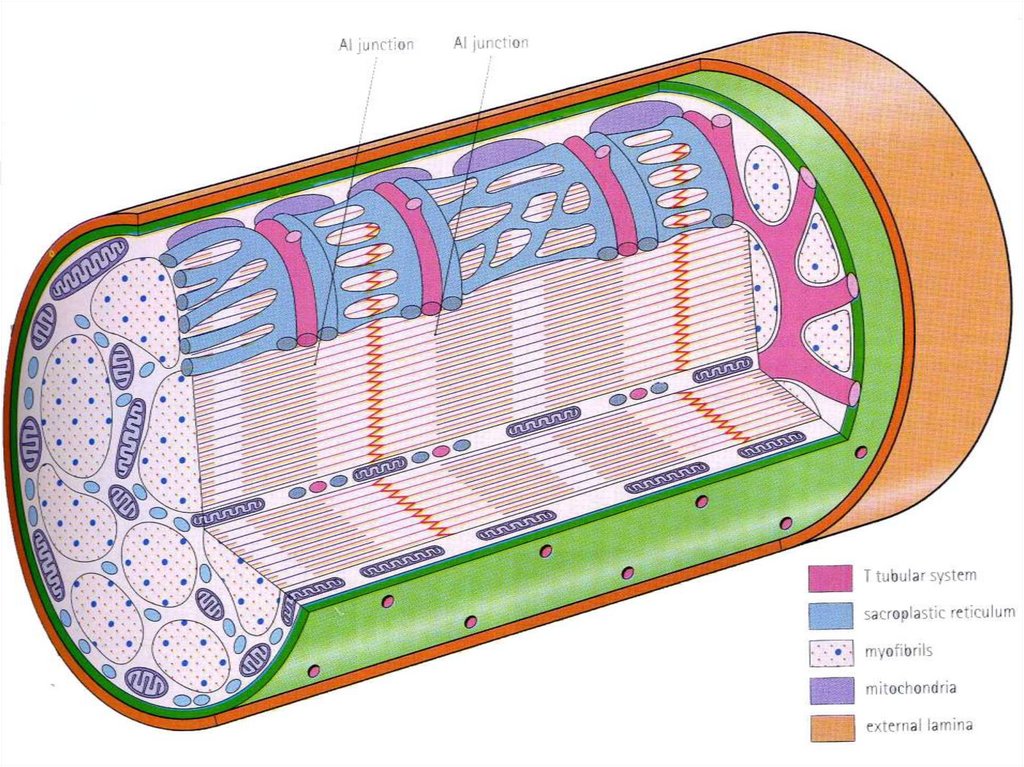
Т -системыинвагинации на уровне
границы между А-и Iдисками дважды в
каждом саркомере.
Они связаны с
терминальными
цистернами СПС –
образуя триады
МИТОХОНДРИИ между
миофибриллами
САРКОЛЕММА БМ +
Ретикулярные волокна
29.
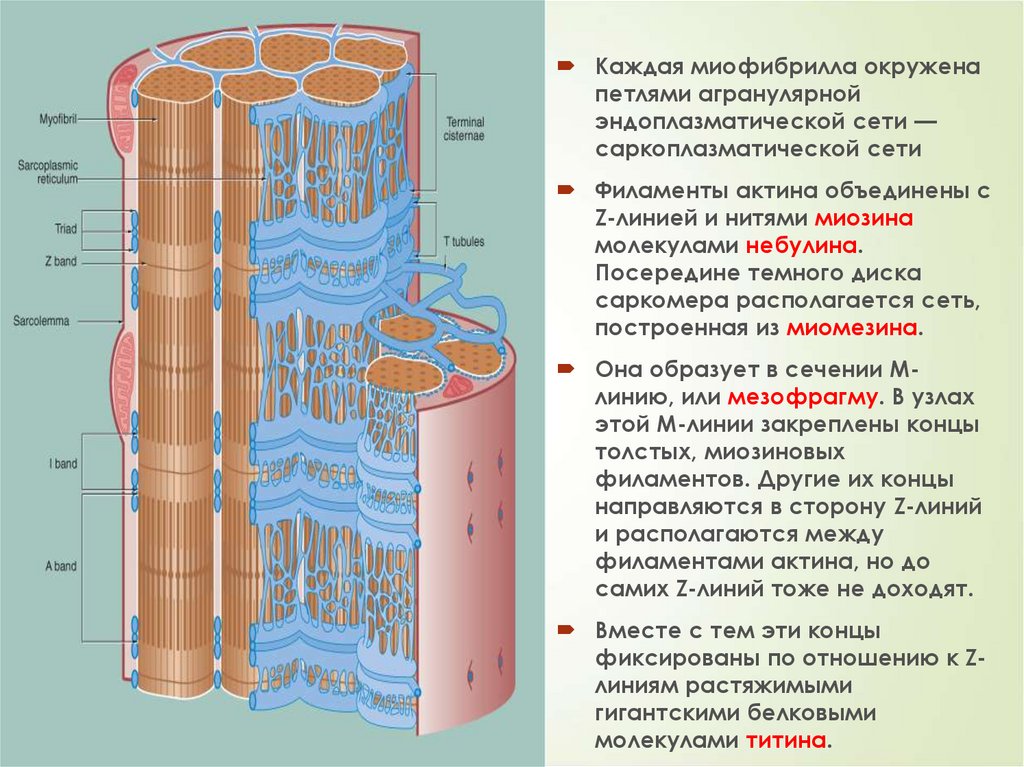
29Каждая миофибрилла окружена
петлями агранулярной
эндоплазматической сети —
саркоплазматической сети
Филаменты актина объединены с
Z-линией и нитями миозина
молекулами небулина.
Посередине темного диска
саркомера располагается сеть,
построенная из миомезина.
Она образует в сечении Млинию, или мезофрагму. В узлах
этой М-линии закреплены концы
толстых, миозиновых
филаментов. Другие их концы
направляются в сторону Z-линий
и располагаются между
филаментами актина, но до
самих Z-линий тоже не доходят.
Вместе с тем эти концы
фиксированы по отношению к Zлиниям растяжимыми
гигантскими белковыми
молекулами титина.
30.
3031.
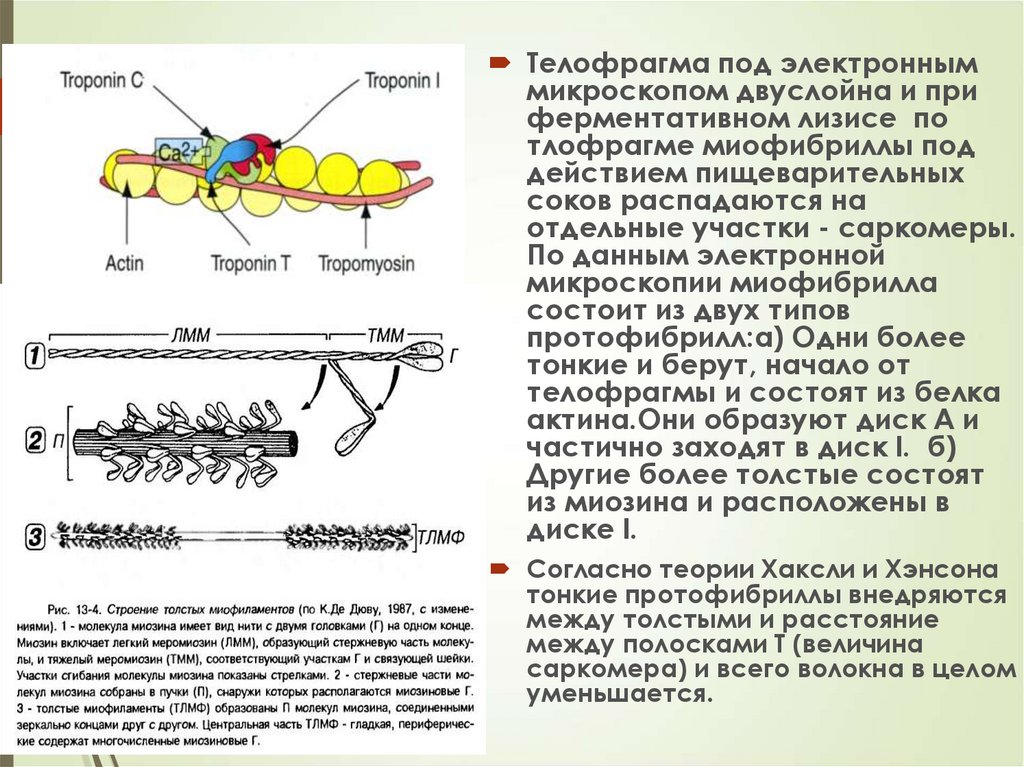
31Телофрагма под электронным
микроскопом двуслойна и при
ферментативном лизисе по
тлофрагме миофибриллы под
действием пищеварительных
соков распадаются на
отдельные участки - саркомеры.
По данным электронной
микроскопии миофибрилла
состоит из двух типов
протофибрилл:а) Одни более
тонкие и берут, начало от
телофрагмы и состоят из белка
актина.Они образуют диск А и
частично заходят в диск I. б)
Другие более толстые состоят
из миозина и расположены в
диске I.
Согласно теории Хаксли и Хэнсона
тонкие протофибриллы внедряются
между толстыми и расстояние
между полосками Т (величина
саркомера) и всего волокна в целом
уменьшается.
32.
3233.
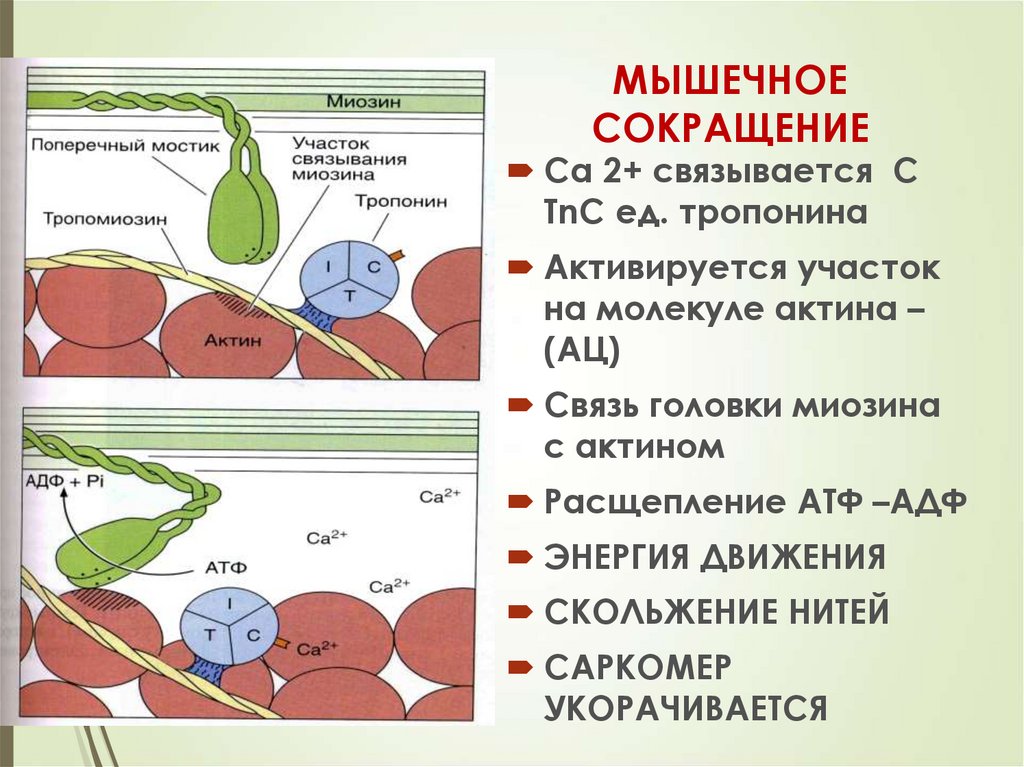
33МЫШЕЧНОЕ
СОКРАЩЕНИЕ
Са 2+ связывается С
ТnC ед. тропонина
Активируется участок
на молекуле актина –
(АЦ)
Связь головки миозина
с актином
Расщепление АТФ –АДФ
ЭНЕРГИЯ ДВИЖЕНИЯ
СКОЛЬЖЕНИЕ НИТЕЙ
САРКОМЕР
УКОРАЧИВАЕТСЯ
34.
3435.
35Белки Опорного Аппарата
и ДИСТРОФИЯ ДЮШЕНА
Белок дистрофин белок одним
концом прикрепляющийся к
актиновым филламентам а другим к
гликопротеинам сарколеммы и Б.М.
Костамеры –кольца из белка
винкулина которые охватывают
волокно и соединяют сарколемму и I
диски
Тетин белок связывающий Z и М линии
препятствует перерастяжению
мышцы
Небулин расположен вдоль тонких
филламентов и
Обеспечивает механическую
стабилизацию
36.
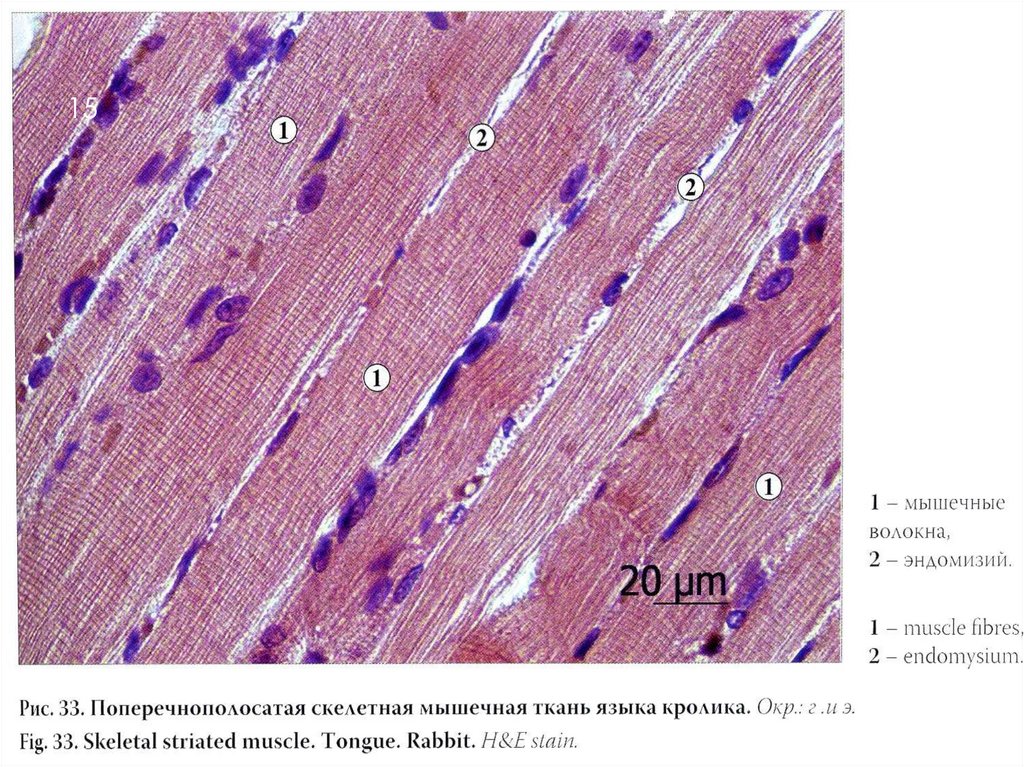
СТРОЕНИЕ МЫШЦЫ – ОРГАННЫЙ УРОВЕНЬ36
Между мышечными волокнами находятся
тонкие прослойки рыхлой волокнистой
соединительной ткани — эндомизий
Более толстые прослойки рыхлой
соединительной ткани окружают по
нескольку мышечных волокон, образуя
перимизий и разделяя мышцу на пучки.
Несколько пучков объединяются в более
крупные группы, разделенные более
толстыми соединительнотканными
прослойками. Соединительную ткань,
окружающую поверхность мышцы,
называют эпимизием
37.
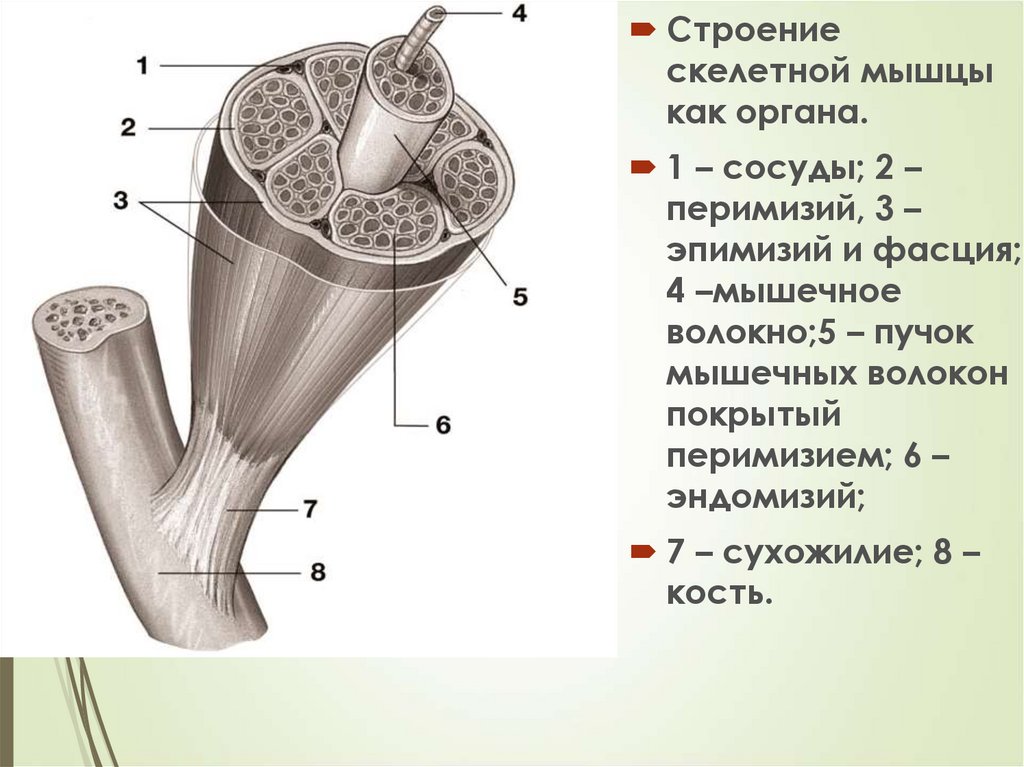
37Строение
скелетной мышцы
как органа.
1 – сосуды; 2 –
перимизий, 3 –
эпимизий и фасция;
4 –мышечное
волокно;5 – пучок
мышечных волокон
покрытый
перимизием; 6 –
эндомизий;
7 – сухожилие; 8 –
кость.
38.
38ВАСКУЛЯРИЗАЦИЯ И ИНЕРВАЦИЯ
Васкуляризация. Артерии вступают в
мышцу и распространяются по
прослойкам соединительной ткани,
постепенно истончаясь. Ветви 5—6-го
порядка образуют в перимизии
артериолы.
В эндомизии расположены капилляры.
Они идут вдоль мышечных волокон,
анастомозируя друг с другом. Венулы,
вены и лимфатические сосуды проходят
рядом с приносящими сосудами.
Иннервация. В мышцах выявлены
миелинизированные эфферентные
(двигательные),
афферентные (чувствительные), а также
немиелинизированные вегетативные
нервные волокна.
39.
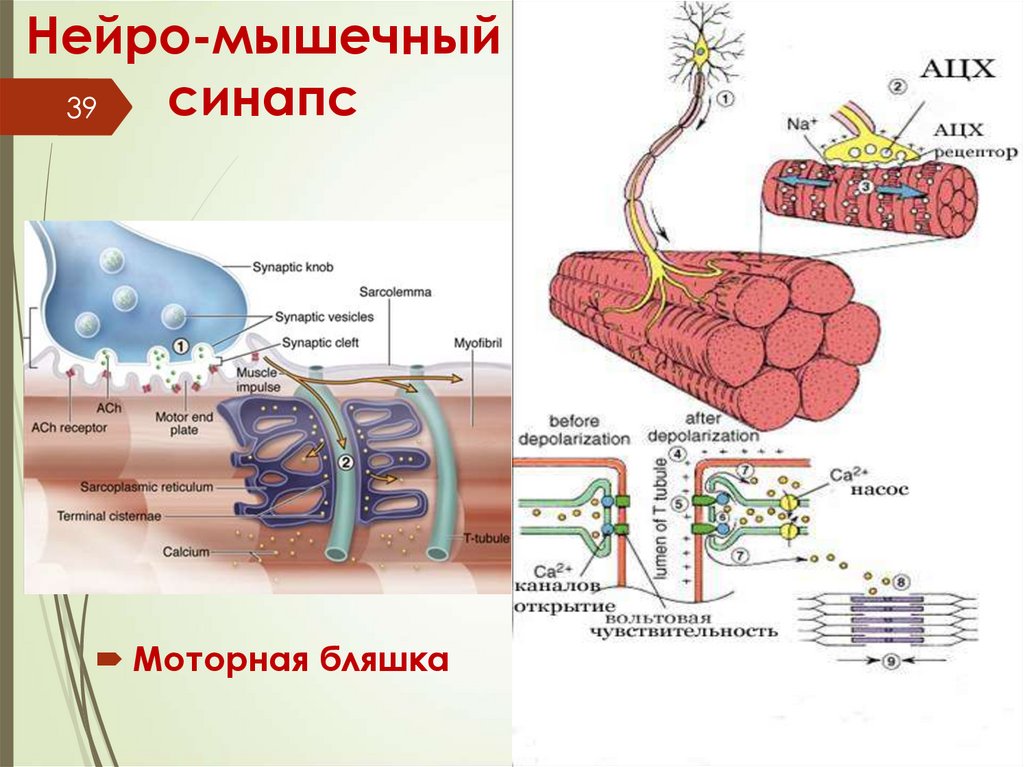
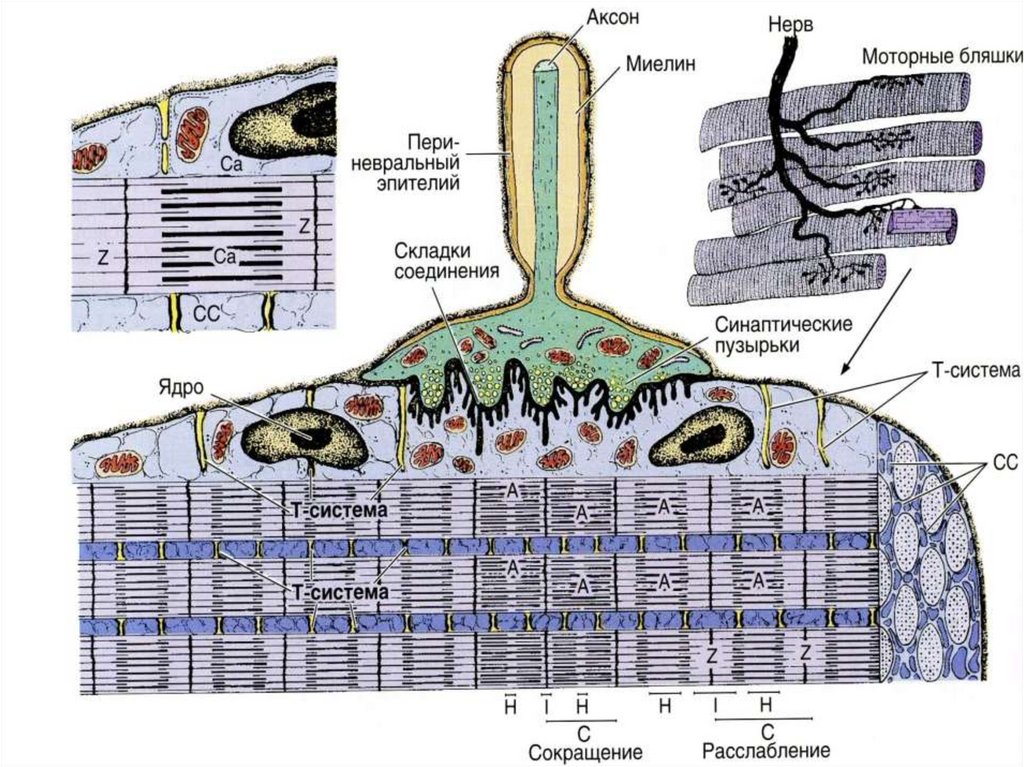
Нейро-мышечныйсинапс
39
Моторная бляшка
40.
40Отросток нервной клетки, приносящий
импульс от моторного нейрона
спинного мозга, ветвится в перимизии.
Каждая его ветвь проникает сквозь
базальную мембрану, и у поверхности
симпласта на плазмолемме образует
терминали, участвуя в организации так
называемой моторной бляшки, или
нервно-мышечного соединения. При
поступлении нервного импульса из
терминали выделяется ацетилхолин —
медиатор, который вызывает
возбуждающий потенциал действия,
распространяющееся отсюда по
плазмолемме миосимпласта.
41.
4142.
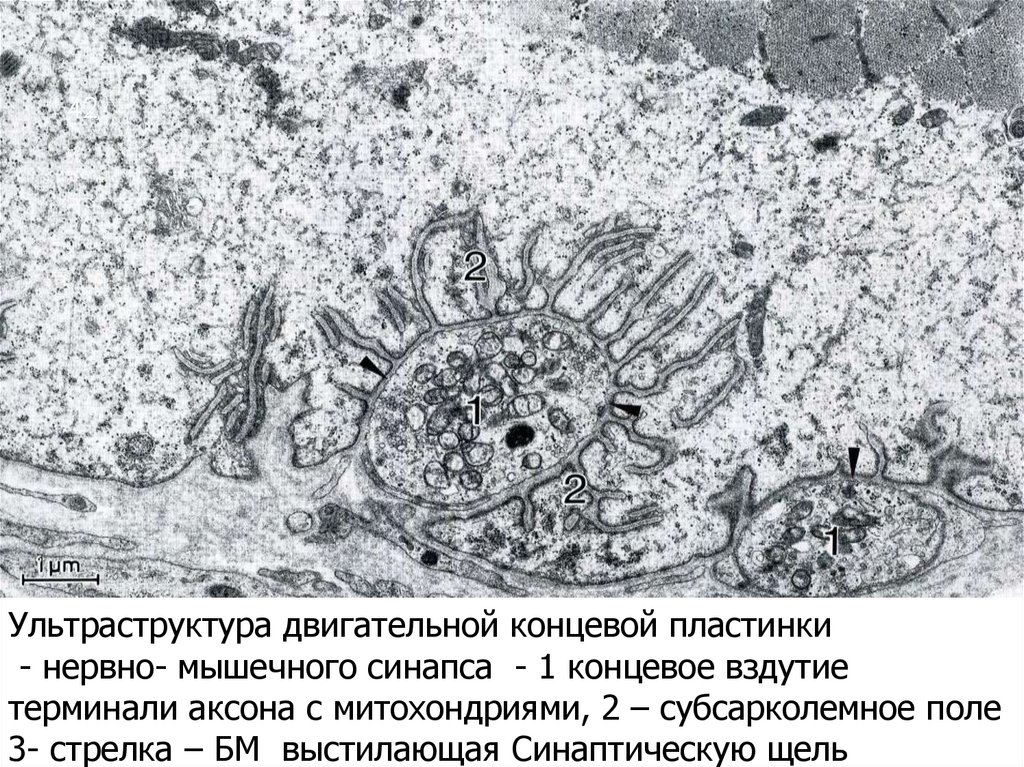
42Ультраструктура двигательной концевой пластинки
- нервно- мышечного синапса - 1 концевое вздутие
терминали аксона с митохондриями, 2 – субсарколемное поле
3- стрелка – БМ выстилающая Синаптическую щель
43.
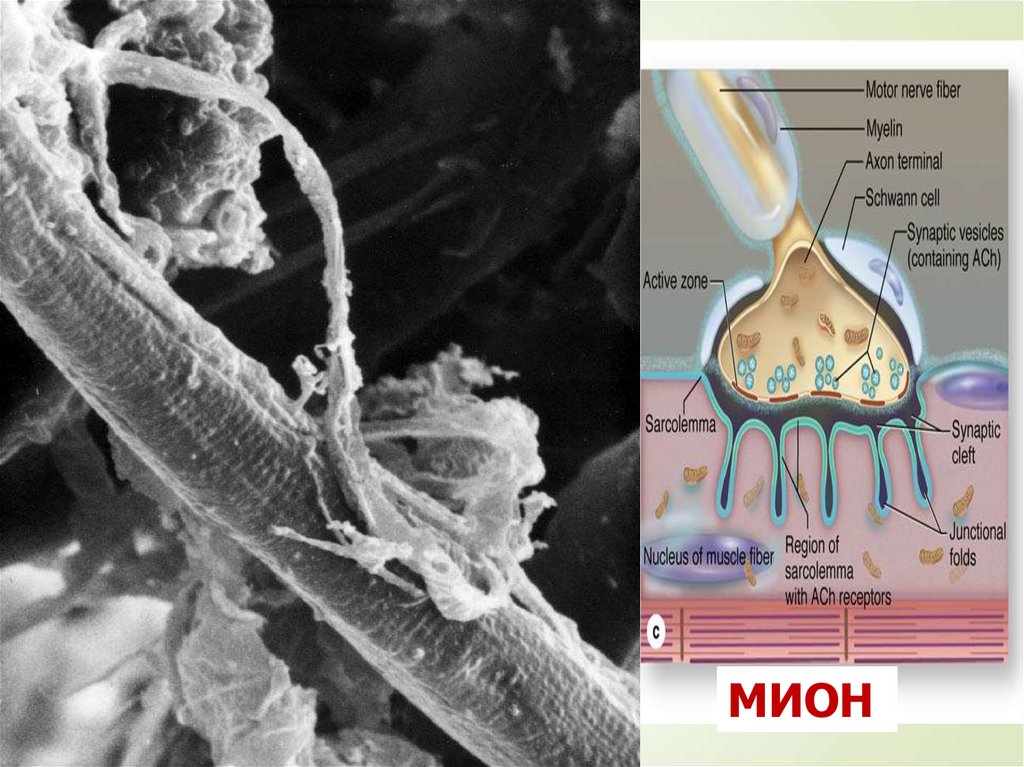
43МИОН
44.
44МИОН
Итак, каждое мышечное волокно
иннервируется самостоятельно и
окружено сетью гемокапилляров,
образуя комплекс, именуемый мионом
Группа же мышечных волокон,
иннервируемых одним мотонейроном,
называется нервно-мышечной
единицей.
Характерно, что мышечные волокна,
принадлежащие к одной нервномышечной единице, лежат не рядом, а
расположены мозаично среди
волокон, относящихся к другим
единицам
45.
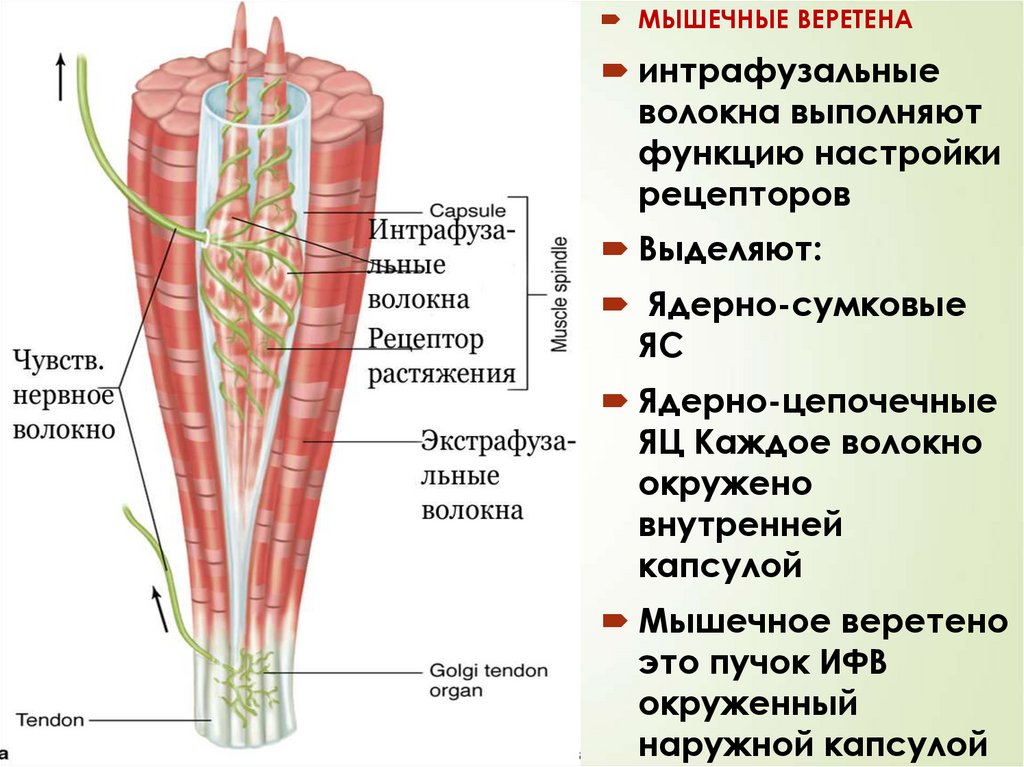
МЫШЕЧНЫЕ ВЕРЕТЕНА45
интрафузальные
волокна выполняют
функцию настройки
рецепторов
Выделяют:
Ядерно-сумковые
ЯС
Ядерно-цепочечные
ЯЦ Каждое волокно
окружено
внутренней
капсулой
Мышечное веретено
это пучок ИФВ
окруженный
наружной капсулой
46.
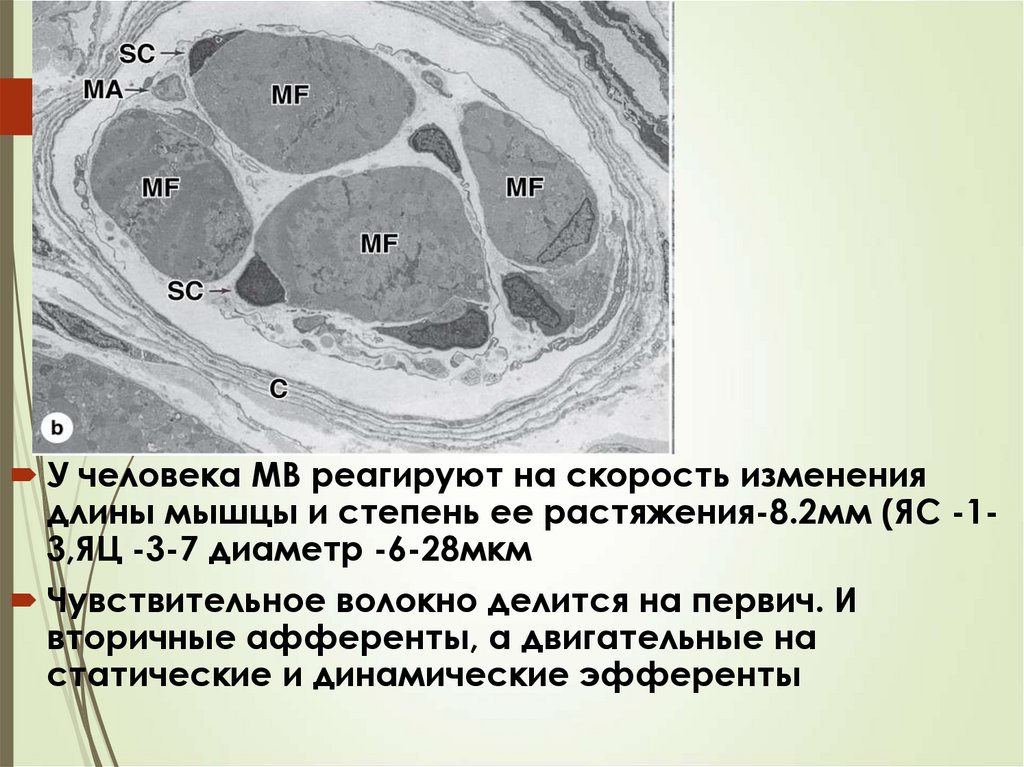
46У человека МВ реагируют на скорость изменения
длины мышцы и степень ее растяжения-8.2мм (ЯС -13,ЯЦ -3-7 диаметр -6-28мкм
Чувствительное волокно делится на первич. И
вторичные афференты, а двигательные на
статические и динамические эфференты
47.
47КРОВЕНОСНЫЕ СОСУДЫ В СКЕЛЕТНОМ
МЫШЕЧНОМ ВОЛОКНЕ. ОКРАСКА ПО ГИМЗЕ +
пластический материал + полляризационный
микроскоп
48.
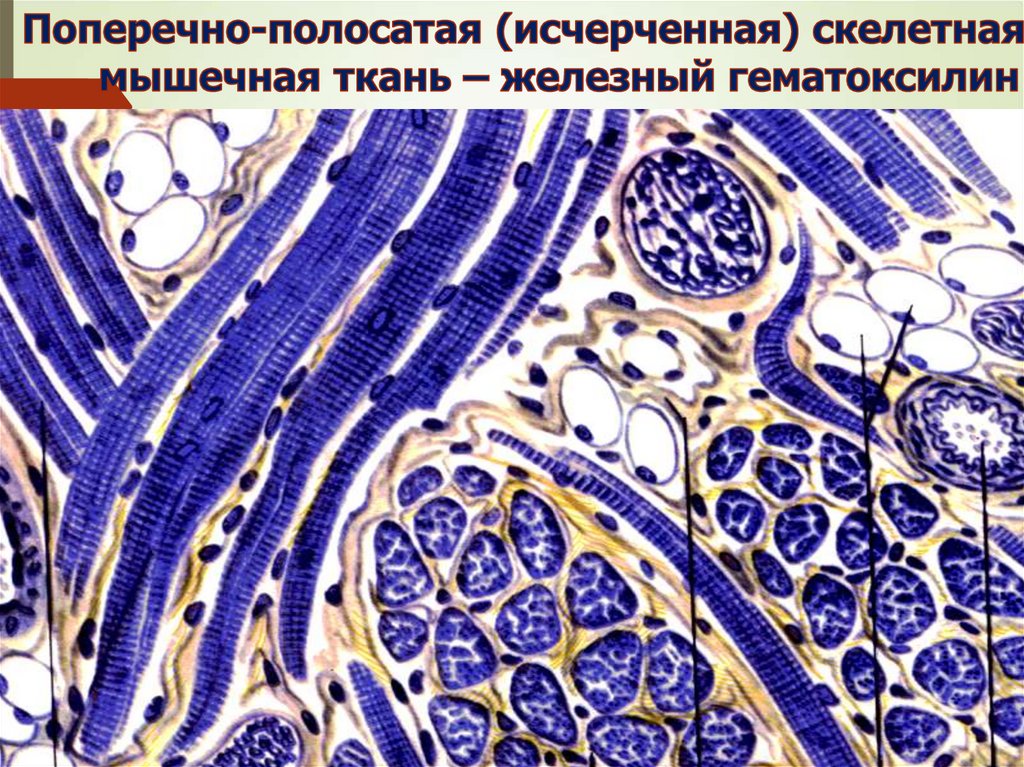
ПОЛЯ КОНГЕЙМА – Fe гематоксилин48
49.
4950.
50СОКРАЩЕНИЕ МЫШЦ
Сокращение мышцы представляет
прерывистый процесс - чем быстрее
восстанавливается затраченная энергия,тем
чаще происходят одно за другим
сокращения
После смерти синтез АТФ прекращается и в
мышцах не оказывается больше
молекул,вызывающих отделение миозина от
актина и сохраняется соединенным
комплекс актомиозина.
Поэтому в течение нескольких часов после
смерти миофиламенты сцеплены между
собой в фиксированном положении
Такое состояние называется трупным
окоченением
51.
ЭКСПЕРИМЕНТЫ СТУДИТСКОГО А.Н.Проведены на крысах,морских свинках,
кроликах, собаках
Оперативное вмешательство: насечки,
полная перерезка, резекция части
мышци, почти полное удаление
Наблюдали полную репаративную
регенерацию скелетных мышц
Студитский
А.Н.
1986
Условие: сохранение натяжения,
инервации и функционального
напряжения
Практика –
Б.М. Карлсон
Видовые
отличия
Регенерация скакательных
мышц(икроножная, подошвенная,
камбаловидная) при пересадке в их ложе
измельченной до состояния фарша
удаленной мышечной ткани – через 1
неделю
Приразвита
денервации
– полоска
У крыс регенерация
лучше
всего, а у собак
соединительной
ткани
хуже всего, нарушается
после ишемии,
атрофии и др.
52.
Понятие о мышечных почках(появляются на концах перерезанных мышц)
При лёгких повреждениях (ожогах)
превалируют процессы с образованием
мышечных почек
При глубоких повреждениях присоединяется
миобластический путь (Клишов А.А. )
Появление миосимпластов и миобластов
есть результат дедифференцировки
поврежденных мышц
Чем глубже дедифференцировка, тем
больше в травмированных мышцах
появляется миобластов
Особенно много при измельчении и
пересадки в зону повреждения
53.
СОХРАНЕНИЕ САРКОЛЕММЫ53
РЕГЕНЕРЦИЯ
КАМБИАЛЬНЫЕ ЭЛЕМЕНТЫ
МИОСАТЕЛЛИТОЦИТЫ
ПО ОКОНЧАНИИ РОСТА –
РЕГЕНЕРАЦИЯ ЗАТУХАЕТ
МЕСТО ПОВРЕЖДЕНИЯ
ФАГОЦИТИРУЮТ МАКРОФАГИ
КОМПЕНСАТОРНАЯ
ГИПЕРТРОФИЯ
ПРОЛИФЕРАЦИЯ
МИОСАТЕЛЛИТОЦИТОВ
ПОВРЕЖДЕННЫЙ КОНЕЦ –
МЫШЕЧНАЯ ПОЧКА
ДЕЛЕНИЕ МСЦ
РЕГЕНЕРАЦИЯ
54.
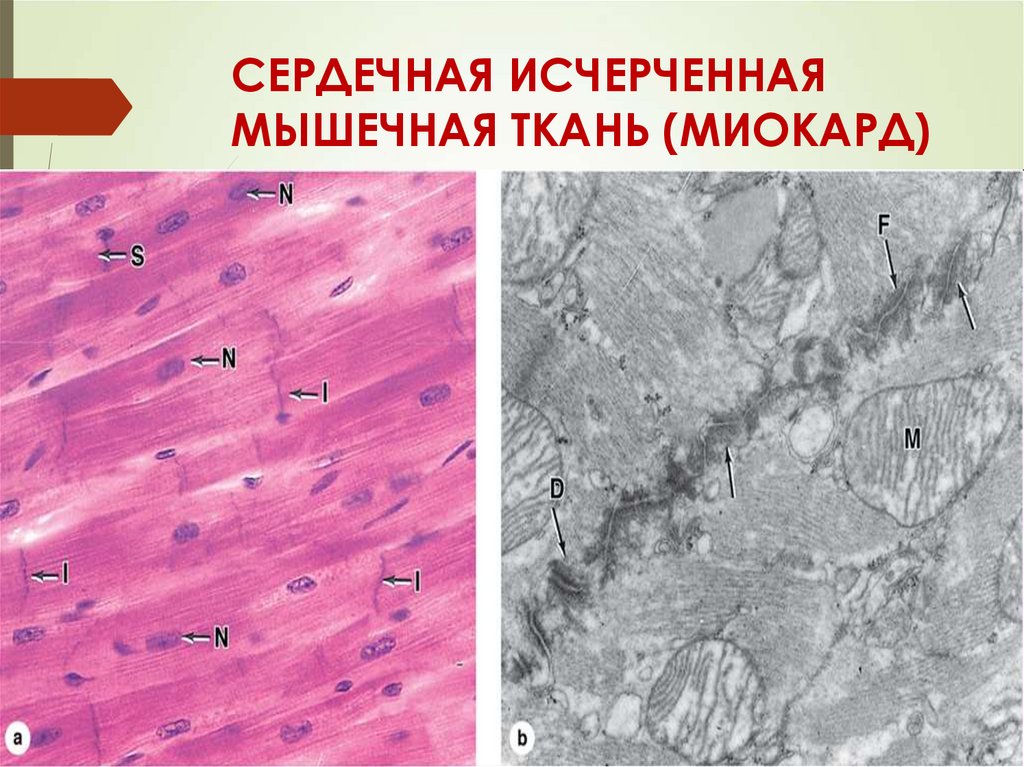
СЕРДЕЧНАЯ ИСЧЕРЧЕННАЯМЫШЕЧНАЯ ТКАНЬ (МИОКАРД)
55.
5556.
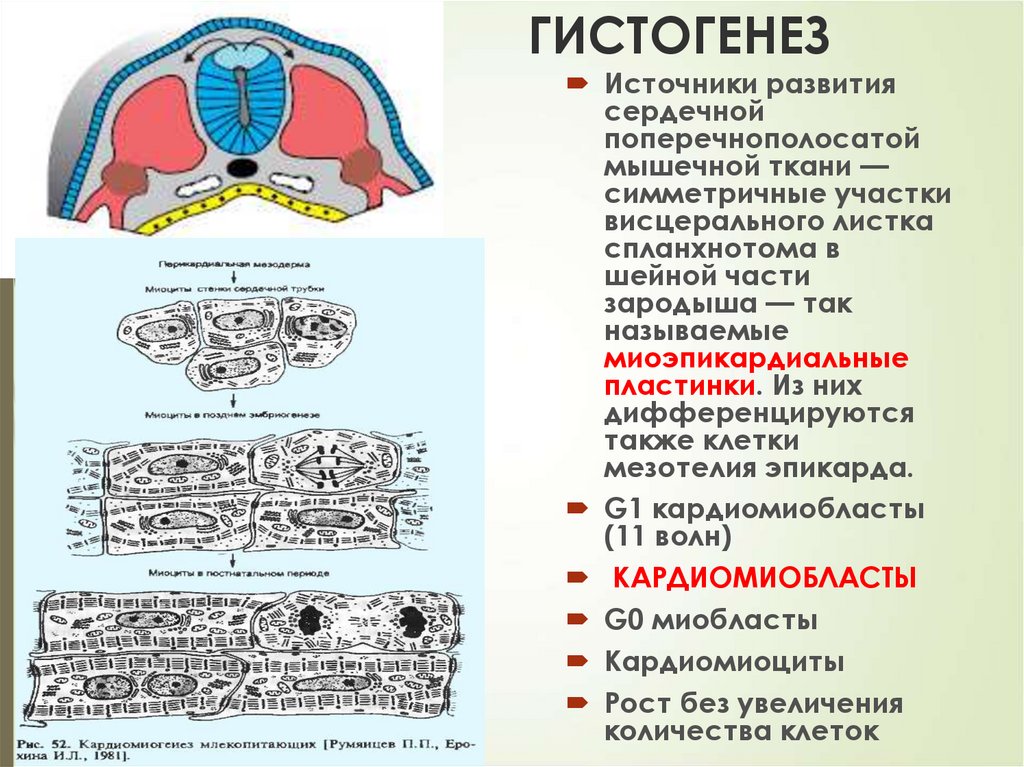
ГИСТОГЕНЕЗ56
Источники развития
сердечной
поперечнополосатой
мышечной ткани —
симметричные участки
висцерального листка
спланхнотома в
шейной части
зародыша — так
называемые
миоэпикардиальные
пластинки. Из них
дифференцируются
также клетки
мезотелия эпикарда.
G1 кардиомиобласты
(11 волн)
КАРДИОМИОБЛАСТЫ
G0 миобласты
Кардиомиоциты
Рост без увеличения
количества клеток
57.
57ДИВЕРГЕНТНАЯ
ДИФЕРЕНЦИРОВКА
КАРДИОМИОЦИТЫ ФОРМИРУЮТ
КОНТАКТЫ «КОНЕЦ В КОНЕЦ»
«КОНЕЦ В БОК» – ФОРМИРУЯ
ФУНКЦИОНАЛЬНЫЙ СИНЦИТИЙ –
СЕРДЕЧНЫЕ МЫШЕЧНЫЕ ВОЛОКНА
-СЕТЕВИДНАЯ СТРУКТУРА
В ДАЛЬНЕЙШЕМ КРОМЕ
СОКРАТИТЕЛЬНО-СЕКРЕТОРНЫХ
ВОЗНИКАЮТ ТЕМНЫЕ СОКРАТИТЕЛЬНЫЕ
СВЕТЛЫЕ ПРОВОДЯЩИЕ
КАРДИОМИОЦИТЫ
СЕРЕТОРНЫЕ ИЛИ ПРЕДСЕРДНЫЕ
КАРДИОМИОЦИТЫ
58.
58ВИДЫ КАРДИОМИОЦИТОВ
рабочие, или типичные, или же
сократительные, кардиомиоциты,
атипичные кардиомиоциты
(пейсмекерные – синусные,
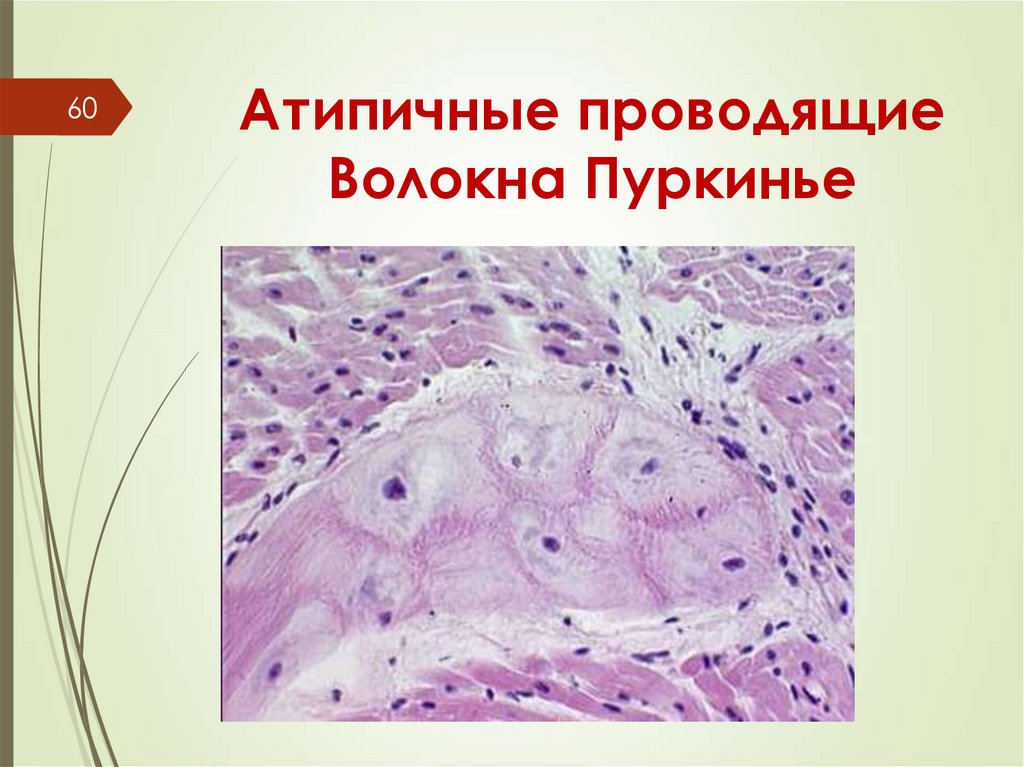
переходные –проводящие
проводящие кардиомиоциты (волокна
Пуркинье , а также
секреторные кардиомиоциты.
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ЕДИНИЦА
- КАРДИОМИОЦИТ
59.
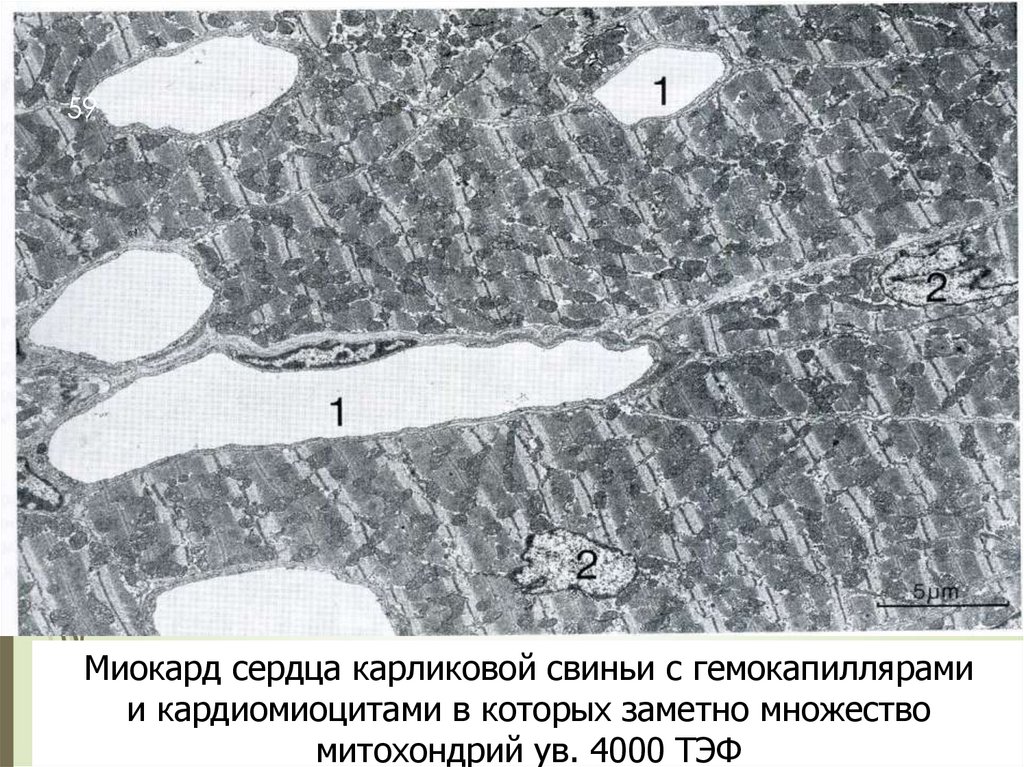
59Миокард сердца карликовой свиньи с гемокапиллярами
и кардиомиоцитами в которых заметно множество
митохондрий ув. 4000 ТЭФ
60.
60Атипичные проводящие
Волокна Пуркинье
61.
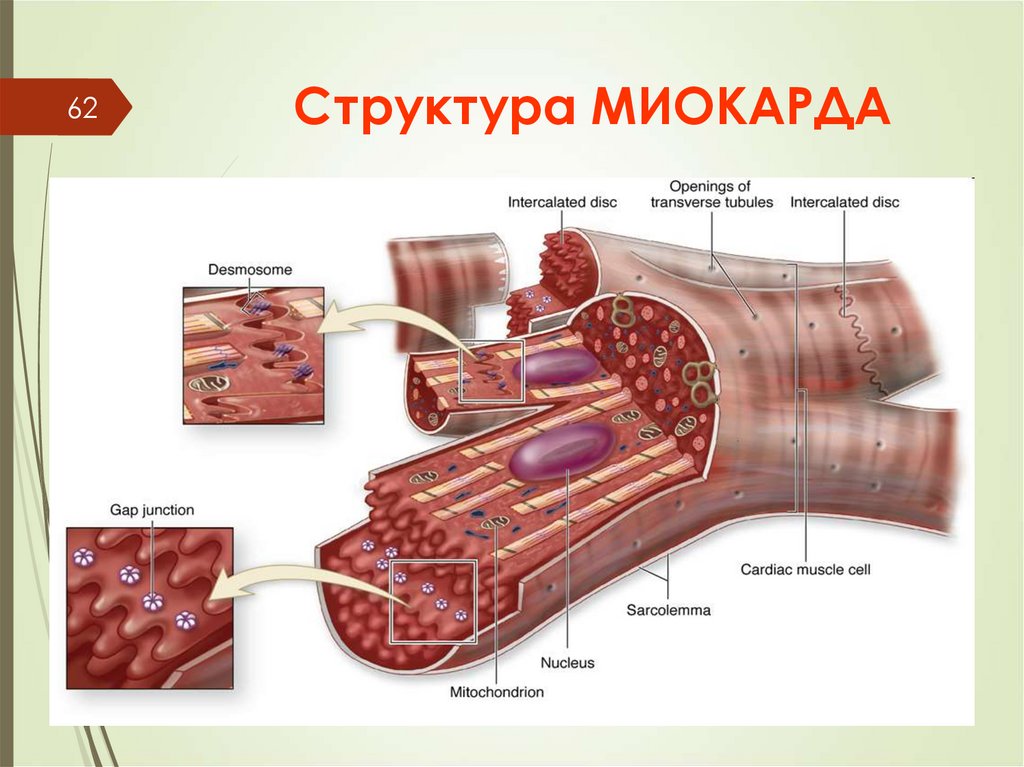
ФУНКЦИОНАЛЬНЫЙ СИНЦИТИЙСократительные кардиомиоциты
имеют
удлиненную (100—150 мкм) форму,
близкую к цилиндрической.
Их концы соединяются друг с другом,
так что цепочки клеток составляют
так называемые функциональные
волокна (толщиной до 20 мкм).
ФУНКЦИОНАЛЬНЫЙ СИНЦИТИЙ
В области контактов клеток
образуются так называемые
вставочные диски.
Кардиомиоциты могут ветвиться и
образуют трехмерную сеть. Их
поверхности покрыты базальной
мембраной, в которую снаружи
вплетаются ретикулярные и
коллагеновые волокна.
62.
62Структура МИОКАРДА
63.
6364.
6465.
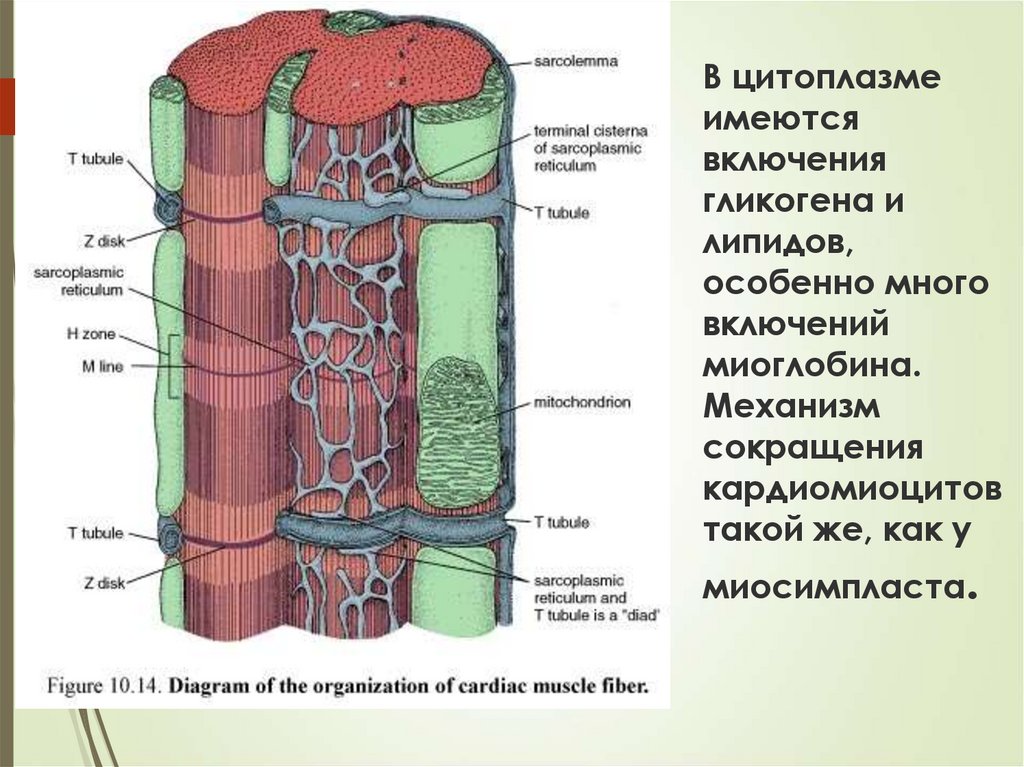
КАРДИОМИОЦИТЫ65
Ядро кардиомиоцита (иногда их два) овальное
и лежит в центральной части клетки. У полюсов
ядра сосредоточены немногочисленные
органеллы общего значения. Миофибриллы
слабо обособлены друг от друга, могут
расщепляться. Их строение аналогично
строению миофибрилл миосимпласта
скелетного мышечного волокна.
От поверхности плазмолеммы в глубь
кардиомиоцита направлены Т-трубочки,
находящиеся на уровне Z-линии.
Их мембраны сближены, контактируют с
мембранами гладкой эндоплазматической
(т.е. саркоплазматической) сети.
Петли последней вытянуты вдоль поверхности
миофибрилл и имеют латеральные утолщения
(L-системы), формирующие вместе с Ттрубочками - диады.
66.
66В цитоплазме
имеются
включения
гликогена и
липидов,
особенно много
включений
миоглобина.
Механизм
сокращения
кардиомиоцитов
такой же, как у
миосимпласта.
67.
6768.
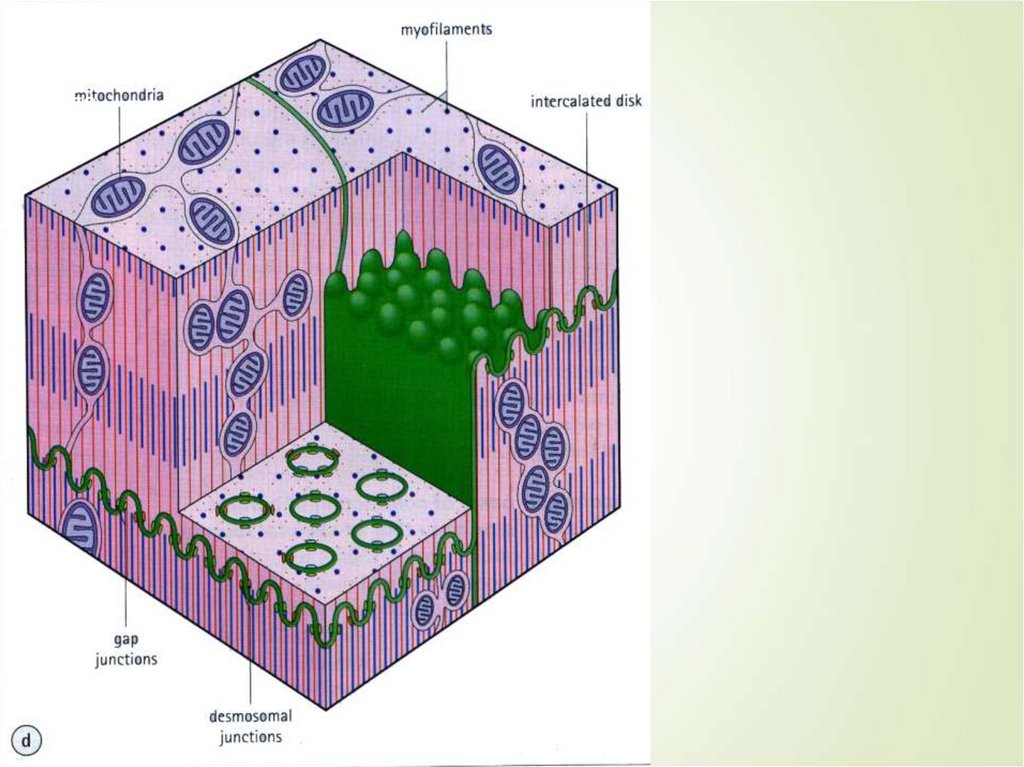
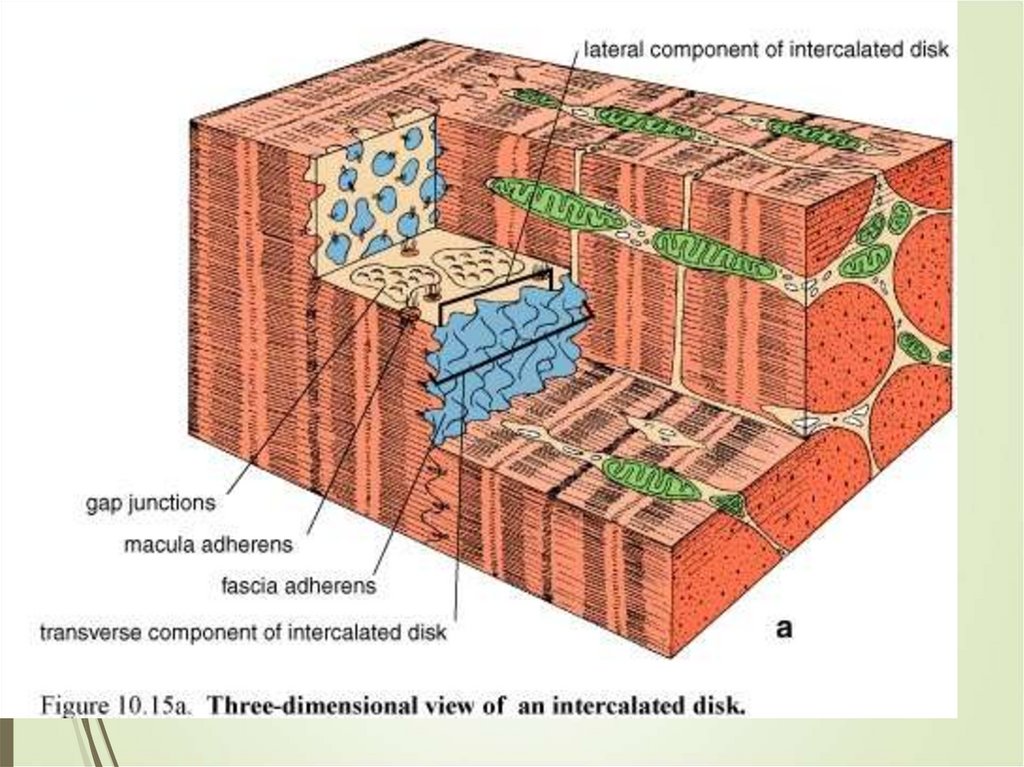
68Кардиомиоциты соединяются друг с другом
своими торцевыми концами. Здесь
образуются так называемые вставочные
диски: эти участки выглядят как тонкие
пластинки на светооптическом уровне
Фактически же концы кардиомиоцитов имеют
неровную поверхность, поэтому выступы одной
клетки входят во впадины другой.
Поперечные участки выступов соседних
клеток соединены друг с другом
интердигитациями и десмосомами. К каждой
десмосоме со стороны цитоплазмы
подходит миофибрилла, закрепляющаяся
концом в десмоплакиновом комплексе.
Таким образом, при сокращении тяга одного
кардиомиоцита передается другому.
Боковые поверхности выступов
кардиомиоцитов объединяются нексусами
(или щелевыми соединениями). Это создает
между ними метаболические связи и
обеспечивает синхронность сокращений
69.
6970.
7071.
7172.
72СЕРЕТОРНЫЕ КАРДИОМИОЦИТЫ
ВЫРАБАТЫВАЮТ ПЕПТИДНЫЕ
ГОРМОНЫ:
КАРДИОДИЛАТИН
АТРИОПЕПТИН
ПРЕДСЕРДНЫЙ НАТРИЙ УРЕТИЧЕСКИЙ
ФАКТОР
Усиливает диурез, понижает
кровянное давление
Усиливает почечный кровоток,
сокращ. Стенки артериол. Усиление
фильтрации и удаление натрия
73.
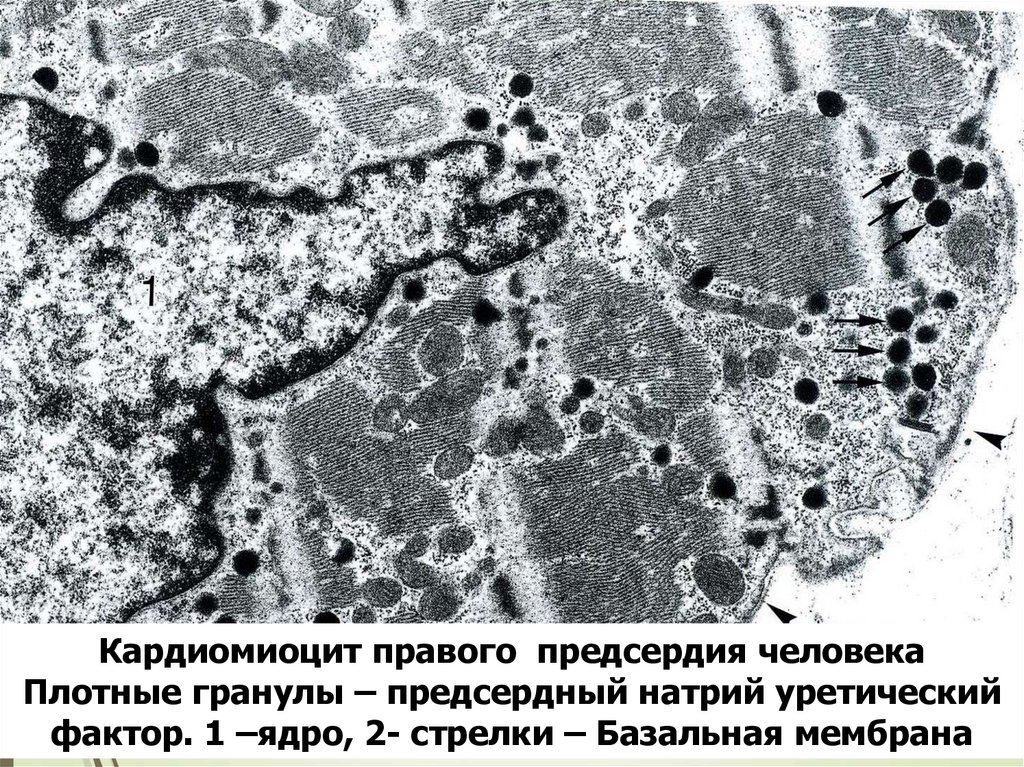
73Кардиомиоцит правого предсердия человека
Плотные гранулы – предсердный натрий уретический
фактор. 1 –ядро, 2- стрелки – Базальная мембрана
74.
74РЕГЕНЕРАЦИЯ
(Румянцев П.П.)
РЕГЕНЕРАЦИЯ ВНУТРИКЛЕТОЧНАЯ ПУТЕМ
ГИПЕРПЛАЗИИ – ПОЛИПЛОИДИЗАЦИИ
АЦИТОКИНЕТИЧЕКИЙ МИТОЗ – МНОГОЯДЕРНОСТЬ
При длительной усиленной работе (например, в
условиях постоянно повышенного
артериального давления крови) происходит
рабочая гипертрофия кардиомиоцитов.
Стволовых клеток или клетокпредшественников в сердечной мышечной
ткани не обнаружено, поэтому погибающие
кардиомиоциты (в частности, при инфаркте
миокарда) не восстанавливаются, а
замещаются элементами соединительной
ткани
75.
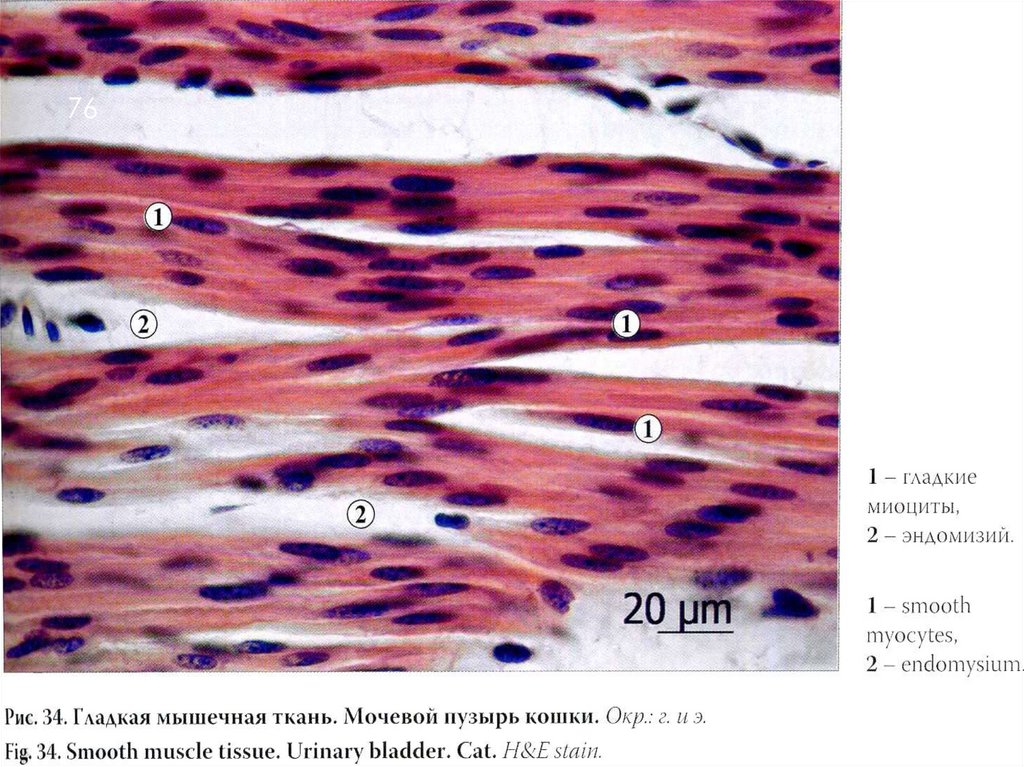
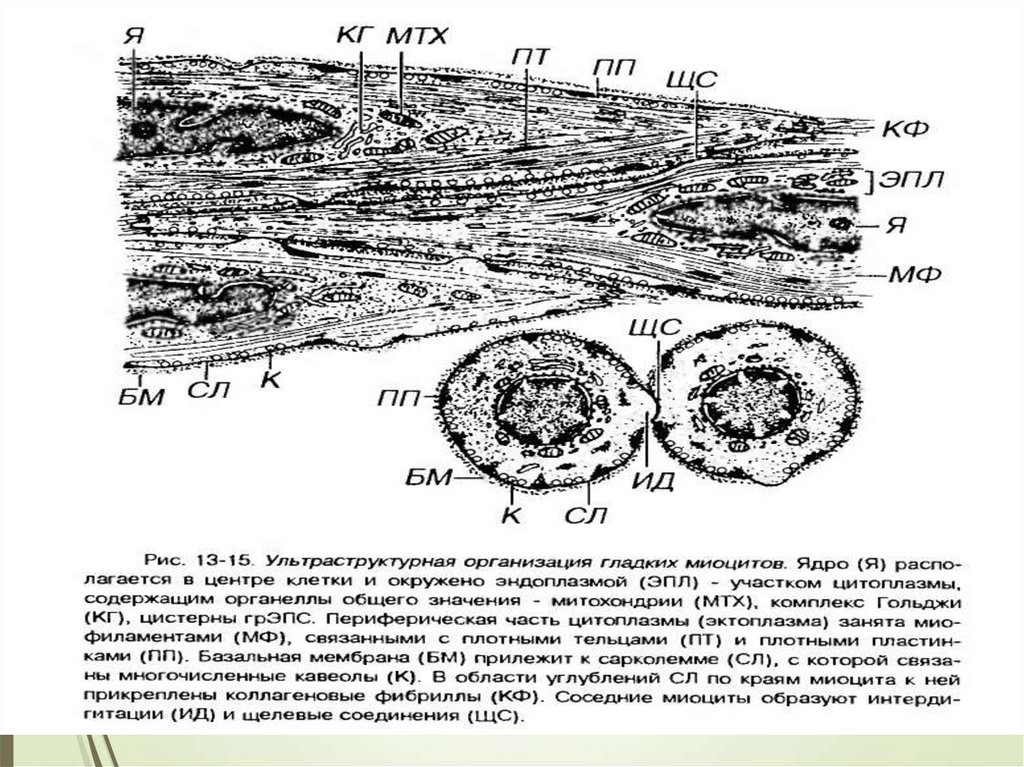
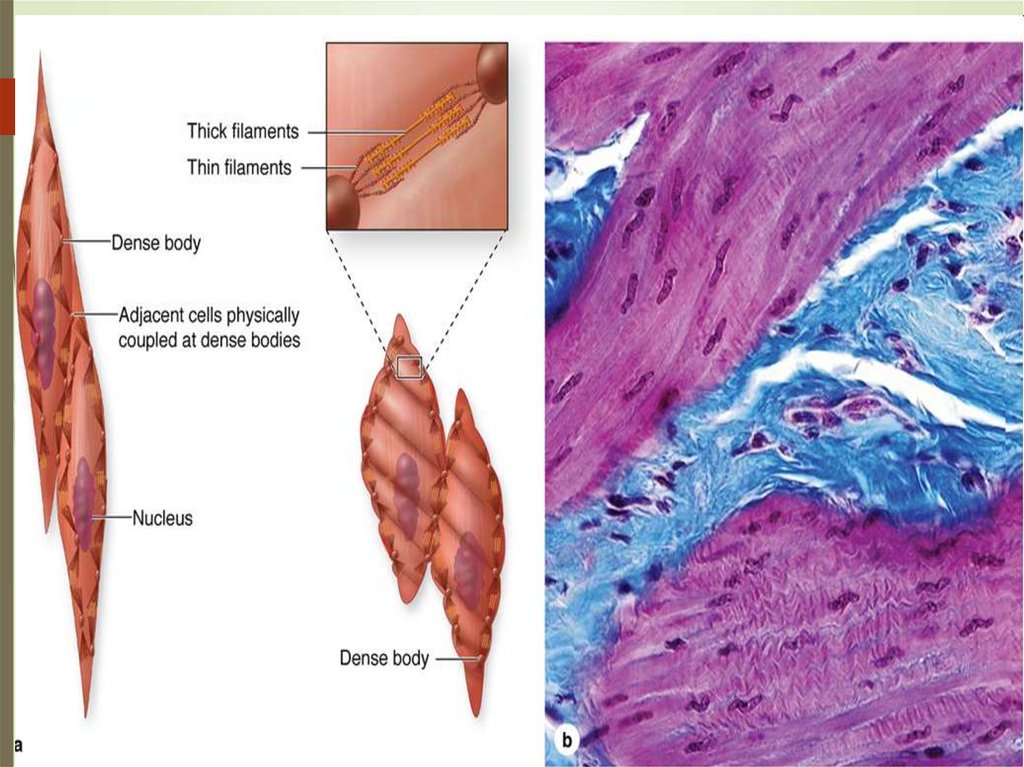
ГЛАДКИЕ МЫШЕЧНЫЕ ТКАНИ75
Структурно-функциональной единицей гладкой, или
неисчерченной, мышечной ткани является гладкомышечная клетка, или гладкий миоцит — это
веретеновидная клетка длиной 20—500 мкм, ширина не
превышает 10 мкм.
Ядро клетки палочковидное, находится в ее
центральной части. Когда миоцит сокращается, его
ядро изгибается и даже закручивается.
Органеллы общего значения, среди которых много
митохондрий, сосредоточены в цитоплазме около
полюсов ядра.
Аппарат Гольджи и гранулярная эндо плазматическая
сеть развиты слабо, что свидетельствует о малой
активности синтетических функций.
Рибосомы в большинстве своем расположены
свободно.
Клетка покрыта саркоплеммой и тонкой базальной
мембраной. Контакты – десмосомы
76.
7677.
77ГИСТОГЕНЕЗ ГЛАДКОЙ МЫШЕЧНОЙ ТКАНИ
ИЗ МЕЗЕНХИМЫ
ВИСЦИРАЛЬНАЯ ГЛАДКАЯ МЫШЕЧНАЯ
ТКАНЬ
СОСУДИСТАЯ ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
ПРОЛИФЕРАТИВНАЯ АКТИВНОСТЬ В
МИОГЕНЕЗЕ СНИЖАЕТСЯ
УВЕЛИЧИВАЕТСЯ КЛ ЦИКЛ, ВЫХОД КЛ ИЗ
ЦИКЛА РЕПРОДУКЦИИ
ДИФФЕРЕНЦИРОВАННЫЕ ГМК
СИНТЕЗИРУЮТ КОЛЛАГЕН И ЭЛАСТИН
78.
7879.
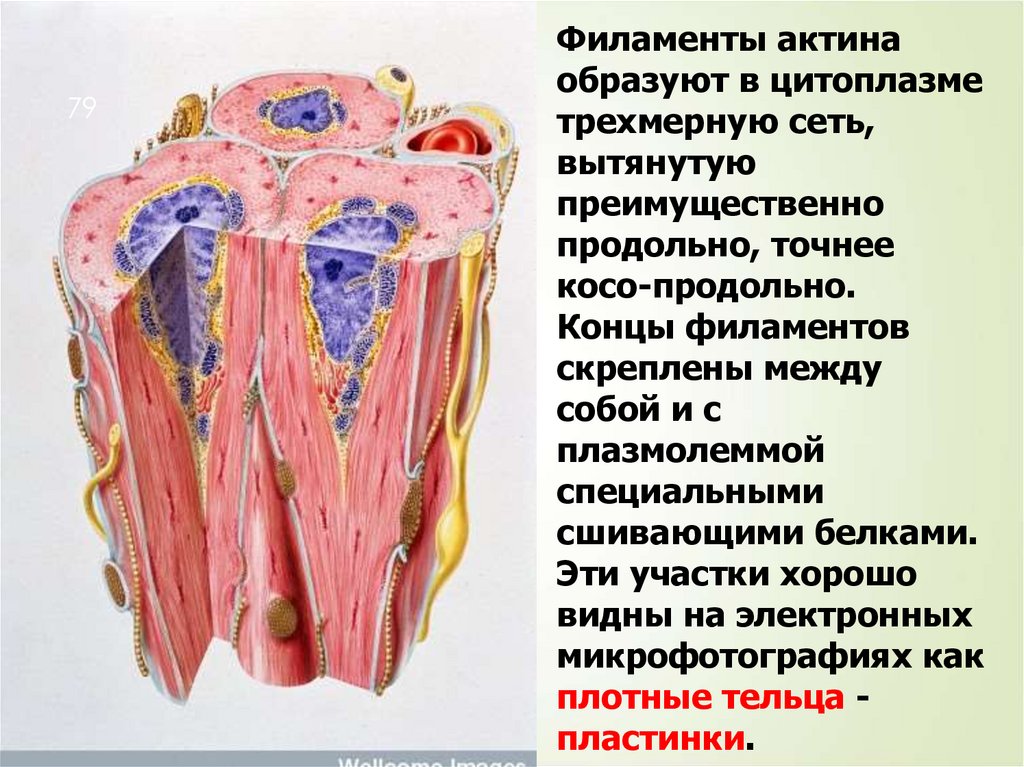
79Филаменты актина
образуют в цитоплазме
трехмерную сеть,
вытянутую
преимущественно
продольно, точнее
косо-продольно.
Концы филаментов
скреплены между
собой и с
плазмолеммой
специальными
сшивающими белками.
Эти участки хорошо
видны на электронных
микрофотографиях как
плотные тельца пластинки.
80.
8081.
8182.
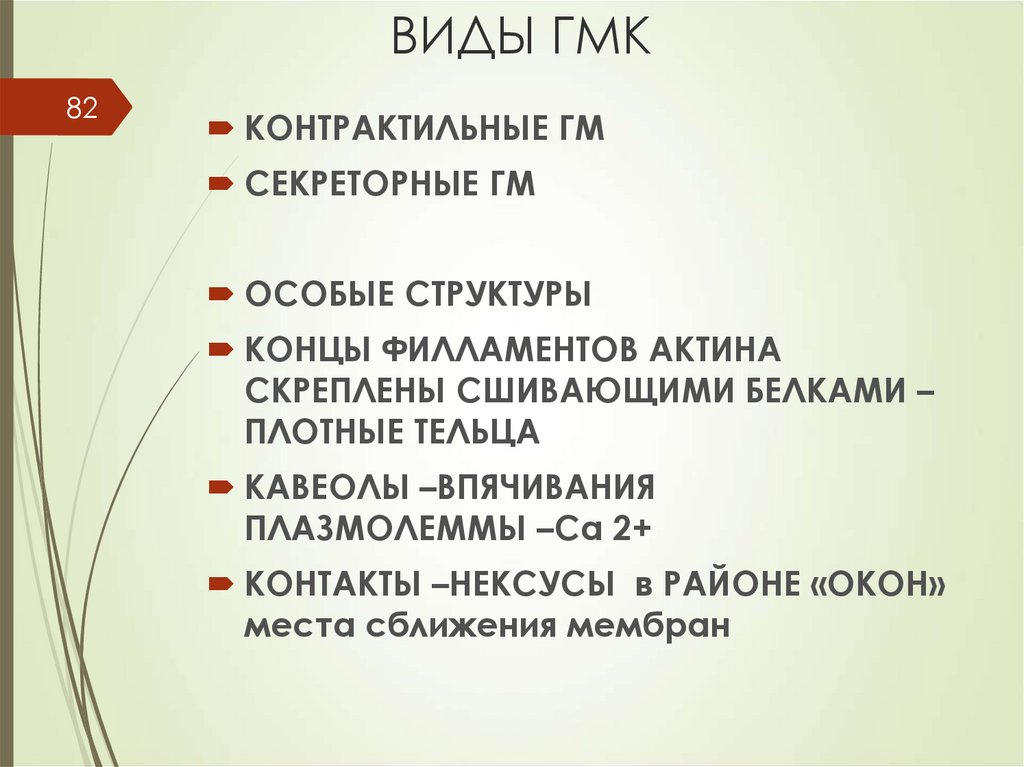
ВИДЫ ГМК82
КОНТРАКТИЛЬНЫЕ ГМ
СЕКРЕТОРНЫЕ ГМ
ОСОБЫЕ СТРУКТУРЫ
КОНЦЫ ФИЛЛАМЕНТОВ АКТИНА
СКРЕПЛЕНЫ СШИВАЮЩИМИ БЕЛКАМИ –
ПЛОТНЫЕ ТЕЛЬЦА
КАВЕОЛЫ –ВПЯЧИВАНИЯ
ПЛАЗМОЛЕММЫ –Са 2+
КОНТАКТЫ –НЕКСУСЫ в РАЙОНЕ «ОКОН»
места сближения мембран
83.
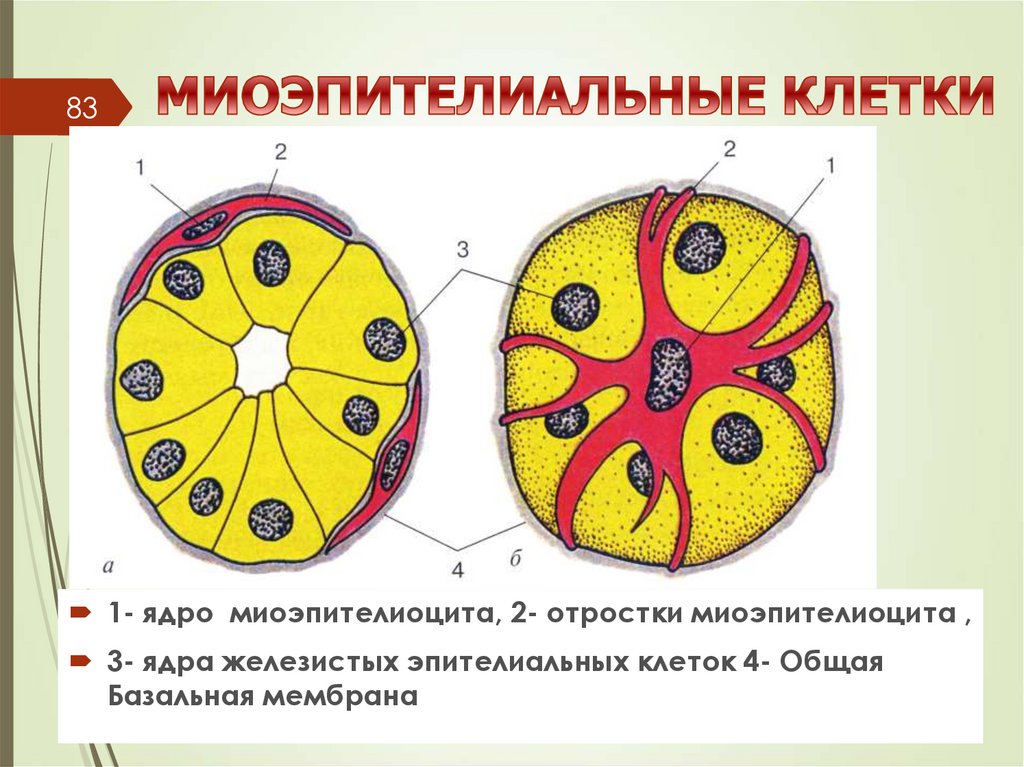
831- ядро миоэпителиоцита, 2- отростки миоэпителиоцита ,
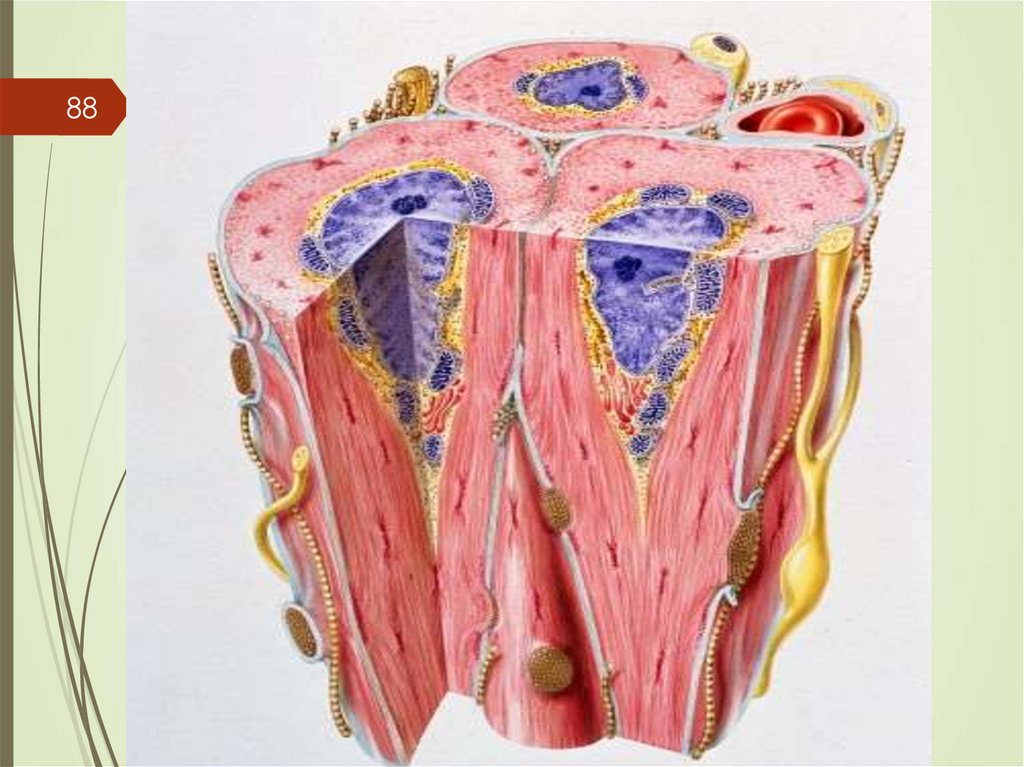
3- ядра железистых эпителиальных клеток 4- Общая
Базальная мембрана
84.
ОТЛИЧИЯ ОТ ДРУГИХ МЫШЦ84
Ионы Са 2+ оказывают влияние не на
актиновые а на миозиновые филаменты
Присутствует цАМФ и особый фермент
киназа , активирующийся комплексом Са +
кальмодулин и регулирующий
фосфорилирование L цепи миозина для
обеспечения скольжения актина
Эстрогены
а прогистерон
уров. цАМФ.
Особый ф-т фосфотаза миозина
прекращает взаимодействие между
филламентами.
85.
85ОСОБЕННОСТЬ
АКТИНОМИОЗИНОВЫЕ
КОМПЛЕКСЫ СУЩЕСТВУЮТ
ТОЛЬКО В ПЕРИОД СОКРАЩЕНИЯ
ПРИ НАЛИЧИИ СВОБОДНЫХ
ИОНОВ Са 2+
86.
86ОТЛИЧИЯ ОТ ДРУГИХ МЫШЦ
Образование мостиков миозина,
которые после дефосфорилирования
не отсоединяются от актина –
поддержание длительного тонуса – без
затрат.
ФИЗИОЛОГИЧЕСКИ путем диффузии
доказано, что функциональным
элементом является –
Пучок гладких миоцитов
87.
8788.
8889.
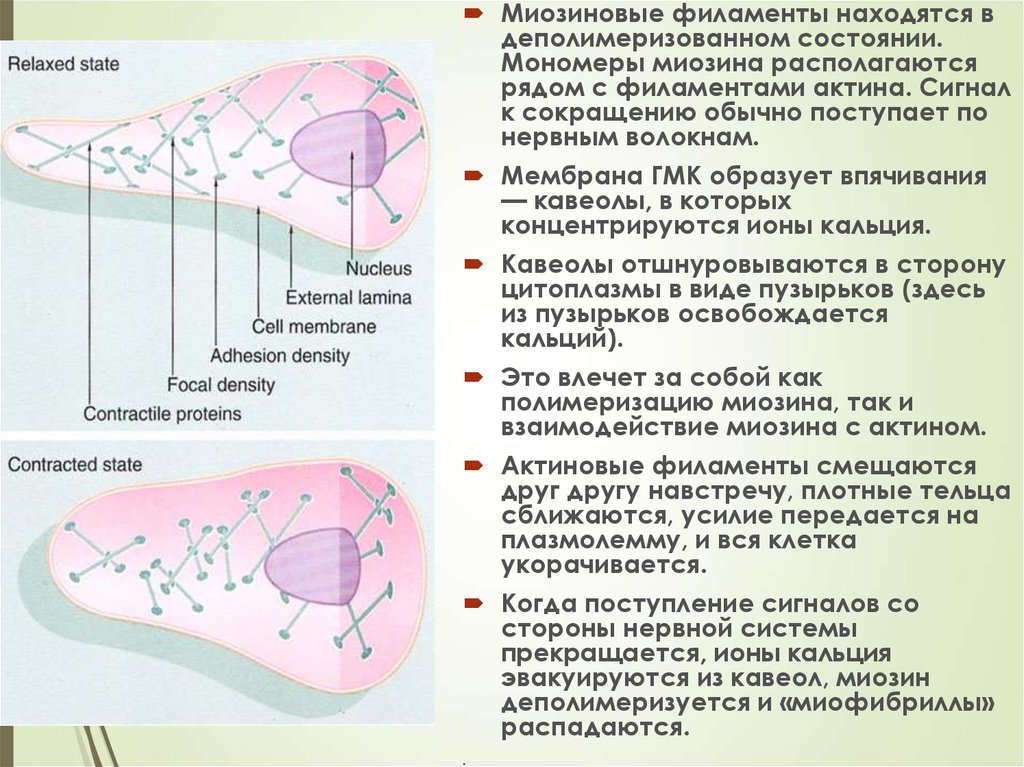
89Миозиновые филаменты находятся в
деполимеризованном состоянии.
Мономеры миозина располагаются
рядом с филаментами актина. Сигнал
к сокращению обычно поступает по
нервным волокнам.
Мембрана ГМК образует впячивания
— кавеолы, в которых
концентрируются ионы кальция.
Кавеолы отшнуровываются в сторону
цитоплазмы в виде пузырьков (здесь
из пузырьков освобождается
кальций).
Это влечет за собой как
полимеризацию миозина, так и
взаимодействие миозина с актином.
Актиновые филаменты смещаются
друг другу навстречу, плотные тельца
сближаются, усилие передается на
плазмолемму, и вся клетка
укорачивается.
Когда поступление сигналов со
стороны нервной системы
прекращается, ионы кальция
эвакуируются из кавеол, миозин
деполимеризуется и «миофибриллы»
распадаются.
.
90.
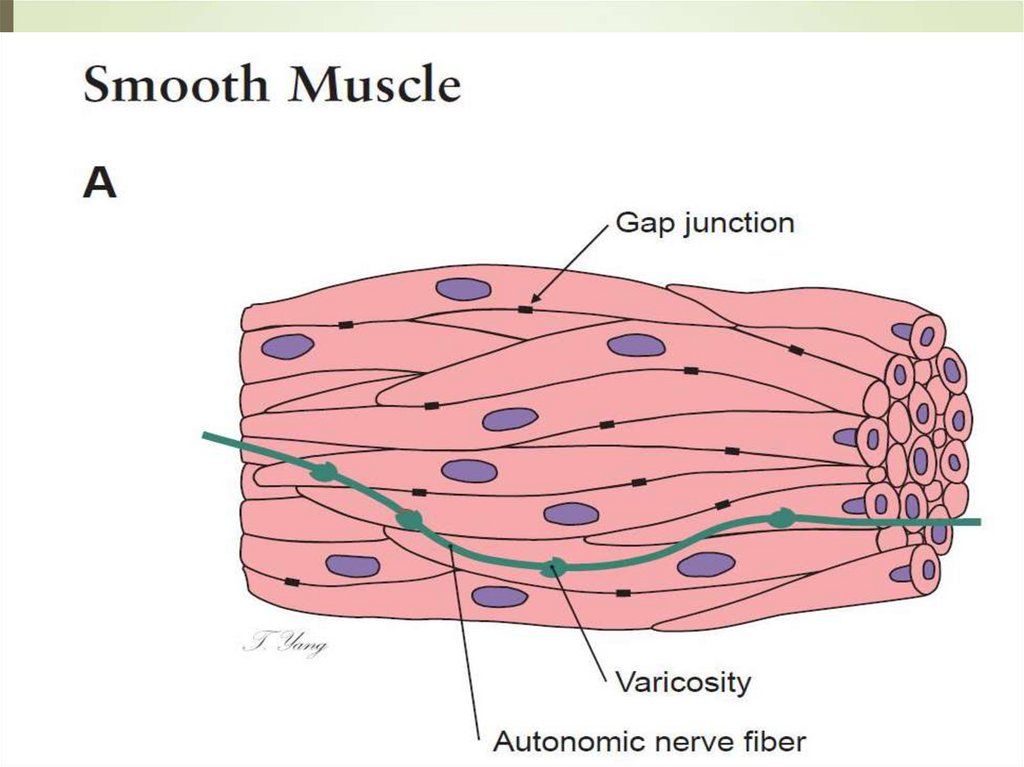
90Таким образом, актино-миозиновые комплексы
существуют в гладких миоцитах только в период
сокращения.
Гладкие миоциты располагаются без заметных
межклеточных пространств и разделены
базальной мембраной. На отдельных участках в
ней образуются «окна», поэтому плазмолеммы
соседних миоцитов сближаются.
Здесь формируются нексусы, и между клетками
возникают не только механические, но и
метаболические связи.
Поверх «чехликов» из базальной мембраны
между миоцитами проходят эластические и
ретикулярные волокна, объединяющие клетки в
единый тканевой комплекс.
Ретикулярные волокна проникают в щели на
концах миоцитов, закрепляются там и передают
усилие сокращения клетки всему их
объединению.
91.
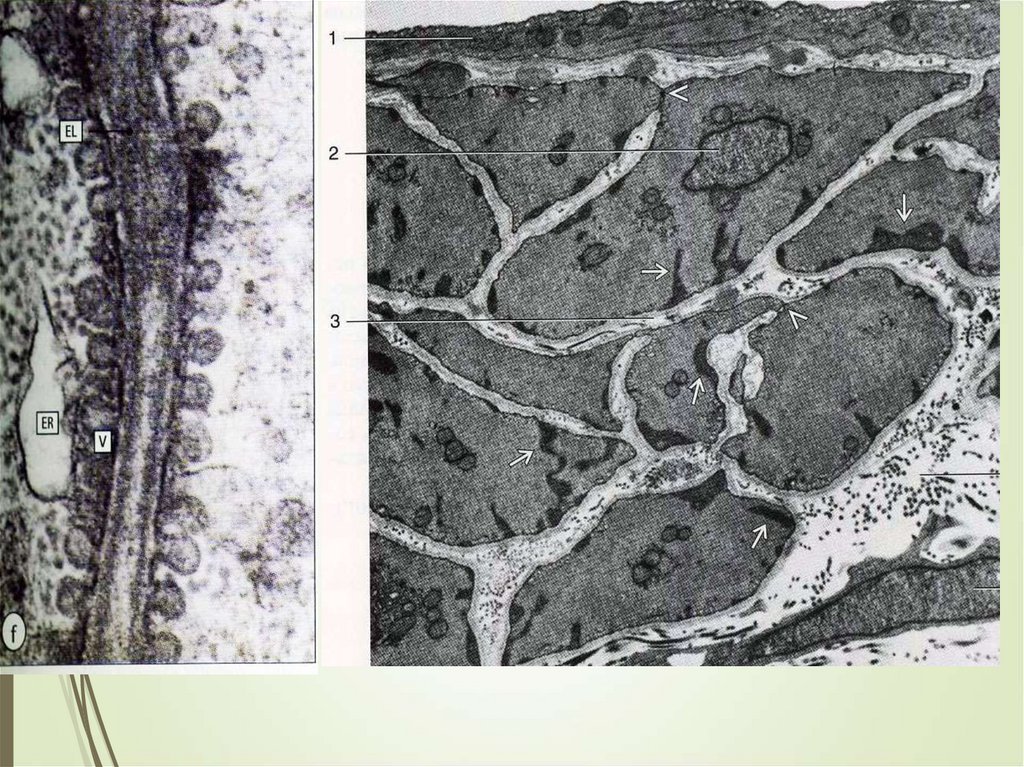
РЕГЕНЕРАЦИЯ91
Физиологическая регенерация
гладкой мышечной ткани проявляется
в условиях повышенных
функциональных нагрузок. Наиболее
отчетливо это видно в мышечной
оболочке матки при беременности.
Такая регенерация осуществляется
не столько на тканевом, сколько на
клеточном уровне: миоциты растут, в
цитоплазме активизируются
синтетические процессы, количество
миофиламентов увеличивается
(рабочая гипертрофия клеток).
Не исключена, однако, и
пролиферация клеток (т.е.
гиперплазия).
92.
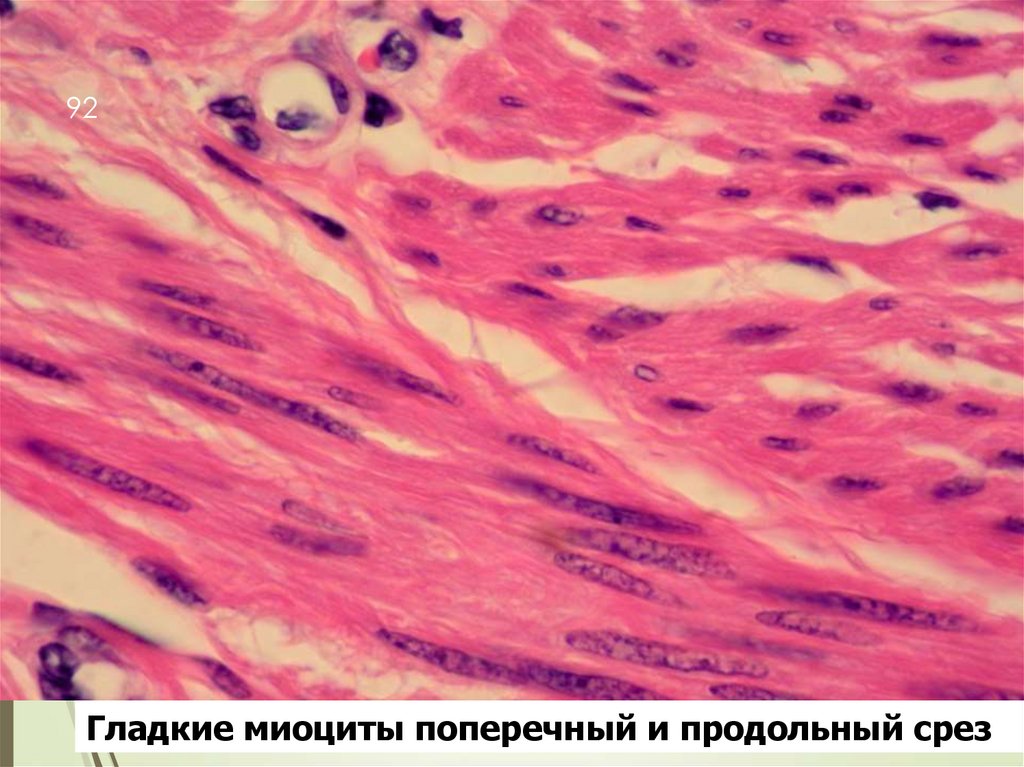
92Гладкие миоциты поперечный и продольный срез
93.
БЛАГОДАРЮЗА ВНИМАНИЕ!