




































 biology
biologySimilar presentations:










Биотехнология: методы молекулярной генетики
1.
БИОТЕХНОЛОГИЯ: МЕТОДЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИШабанова Екатерина Александровна
научный сотрудник ФГБУ «ВНИИЛГИСбиотех»
педагог д/о ГАНОУ «Региональный центр «Орион»
2.
ГЕНЕТИЧЕСКАЯ ИНФОРМАЦИЯВоспроизведение себе подобных является одним из фундаментальных свойств живого. Благодаря этому
явлению существует сходство не только между организмами, но и между отдельными клетками одного
организма. Материальной основой этого сходства является передача генетической информации. Эта
информация зашифрована в последовательности элементов сложных органических молекул – нуклеиновых
кислот ДНК и РНК. Все признаки и свойства клеток и организмов реализуются благодаря белкам, структура
которых закодирована в последовательностях нуклеиновых кислот. Каждая аминокислота (составной элемент
белка) закодирована в ДНК тремя идущими подряд в цепочке нуклеотидами — триплетом.
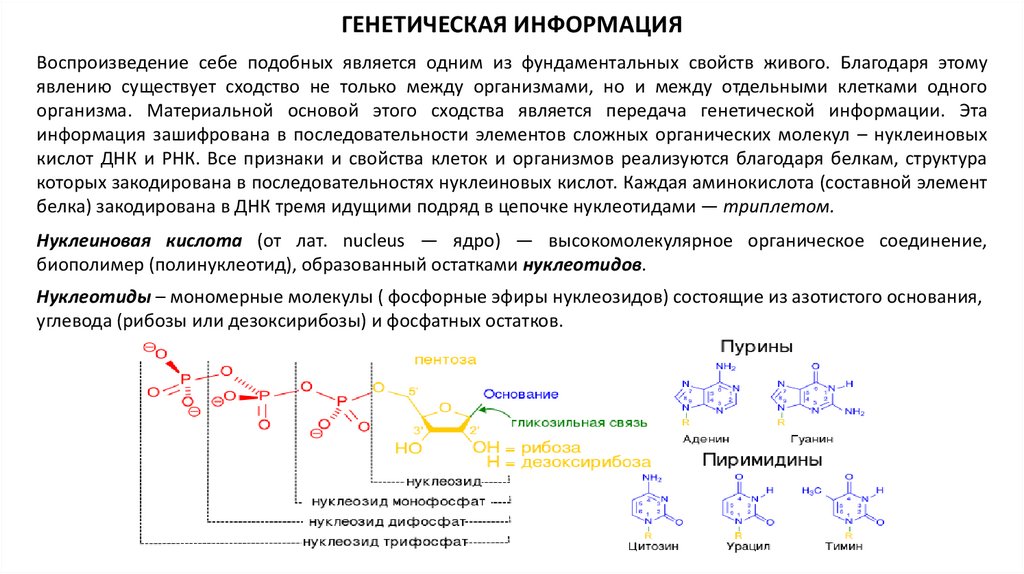
Нуклеиновая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое соединение,
биополимер (полинуклеотид), образованный остатками нуклеотидов.
Нуклеотиды – мономерные молекулы ( фосфорные эфиры нуклеозидов) состоящие из азотистого основания,
углевода (рибозы или дезоксирибозы) и фосфатных остатков.
3.
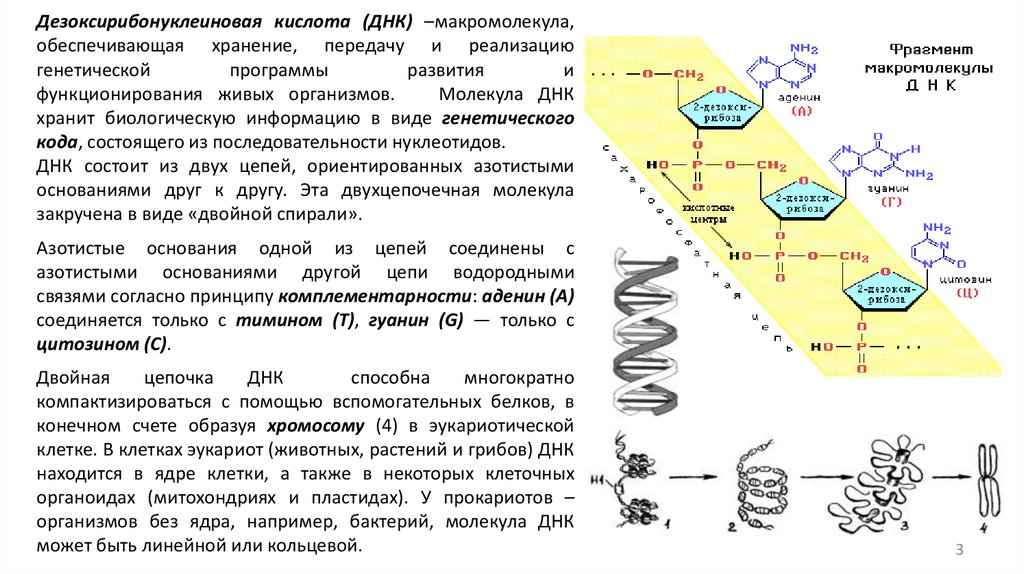
Дезоксирибонуклеиновая кислота (ДНК) –макромолекула,обеспечивающая хранение, передачу и реализацию
генетической
программы
развития
и
функционирования живых организмов.
Молекула ДНК
хранит биологическую информацию в виде генетического
кода, состоящего из последовательности нуклеотидов.
ДНК состоит из двух цепей, ориентированных азотистыми
основаниями друг к другу. Эта двухцепочечная молекула
закручена в виде «двойной спирали».
Азотистые основания одной из цепей соединены с
азотистыми основаниями другой цепи водородными
связями согласно принципу комплементарности: аденин (A)
соединяется только с тимином (T), гуанин (G) — только с
цитозином (C).
Двойная
цепочка
ДНК
способна
многократно
компактизироваться с помощью вспомогательных белков, в
конечном счете образуя хромосому (4) в эукариотической
клетке. В клетках эукариот (животных, растений и грибов) ДНК
находится в ядре клетки, а также в некоторых клеточных
органоидах (митохондриях и пластидах). У прокариотов –
организмов без ядра, например, бактерий, молекула ДНК
может быть линейной или кольцевой.
3
4.
Рибонуклеиноваякислота
(РНК)
состоит из длинной цепи, в которой
каждое звено называется нуклеотидом.
Каждый нуклеотид состоит
из
азотистого основания, сахара рибозы (в
обличии от ДНК, которая содержит
дезоксирибозу) и фосфатной группы.
Последовательность
нуклеотидов
позволяет
РНК
кодировать
генетическую
информацию.
РНК
является одноцепочечной молекулой.
В отличии от ДНК в РНК азотистое
основание
тимин
заменено
на
азотистое основание урацил (U),
которое способно комплементарно
соединяться с аденином (А).
Существует несколько типов РНК: матричная (информационная) РНК служит посредником при передаче
информации, закодированной в ДНК к рибосомам (синтезирующим белки организма), транспортные РНК
(тРНК) и рибосомальные РНК (рРНК), которые участвуют в процессе синтеза белка.
5.
Генетика — наука, изучающая наследственность и изменчивость организмов.Наследственность — способность организмов передавать из поколения в поколение свои
признаки
(особенности
строения,
функций,
развития).
Изменчивость — способность организмов приобретать новые признаки. Наследственность и
изменчивость — два противоположных, но взаимосвязанных свойства организма.
Материальной основой наследственности являются
хромосомы, представляющие собой комплекс
молекулы ДНК (дезоксирибонуклеоновая кислота) с
белками гистонами. Количество хромосом организма
в течение всей жизни постоянно, и, в большинстве
случаев,
число
хромосом
одинаково
у
представителей одного вида. У большинства высших
организмов соматические клетки (клетки тела)
имеют двойной (диплоидный – 2n) набор хромосом,
т.е. каждая хромосома имеет парную, идентичную по
строению и форме гомологичную хромосому.
Половые клетки (гаметы) имеют одинарный
(гаплоидный - n) набор хромосом. При слиянии двух
половых клеток (оплодотворении) у потомства
восстанавливается двойной набор хромосом.
6.
У ряда организмов выделяют соматические хромосомы(аутосомы) и половые хромосомы. Соотношение половых
хромосом опрелеляют пол организма. Например, у
человека в соматической клетке находится 46 хромосом:
44 аутосомы и 2 половые хромосомы. Женщины имеют
две X-хромосомы, а мужчины – одну X и одну Yхромосому. В половых клетках присутствует только одна
половая хромосома, у женщин – это всегда X, а у мужчин –
либо X, либо Y.
Кариотип — совокупность признаков (число, размеры,
форма и т. д.) полного набора хромосом, присущая
клеткам
данного
биологического
вида
(видовой
кариотип),
данного
организма
(индивидуальный кариотип)
Ген — единица наследственной информации, участок молекулы ДНК, отвечающий за один признак, т. е.
кодирующий информацию о структуре определенной молекулы белка (полипептида).
Аллели — различные состояния одного и того же гена, располагающиеся в определённом локусе (участке)
гомологичных хромосом и определяющие развитие признака. Аллели генов определяют наследственные
признаки организмов, передающиеся от родителей потомству при размножении. Аллельные гены
расположены в одинаковых локусах (идентичных участках) гомологичных хромосом
7.
Соматические клетки содержат двааллеля каждого гена, т.к. имеют
двойной набор хромосом.
В
случае, если в гомологичных
хромосомах аллели одинаковые,
организм называется гомозиготой,
а если разные – гетерозиготой.
Доминантным называется аллель, который подавляет проявление другого аллеля
(рецессивного) у гетерозигот, таким образом определяет проявление признака (фенотип) как у
доминантных гомозигот, так и у гетерозигот. Рецессивный аллель может обеспечить проявление
признака только в если находится в гомозиготном состоянии (в паре с таким же рецессивным
аллелем).
Генотип — совокупность генов одной особи, часто под генотипом понимают одну или несколько
аллельных пар генов, которые отвечают за определенные признаки.
Фенотип – совокупность внешних и внутренних признаков организма, проявляющихся в
результате взатмодействия генотипа организма и окружающей среды.
8.
Родоначальником генетики считается австрийскийученый Грегор Мендель (1822-1884), который
впервые применил гибридологический метод,
результатом его исследований явилось открытие
закономерностей наследования, которые сегодня
известны как законы Менделя. Эти принципы
послужили основой для классической генетики и
впоследствии были объяснены как следствие
молекулярных механизмов наследственности.
В основе гибридологического анализа лежит специально
разработанная система скрещиваний. При записи
скрещиваний употребляются стандартные обозначения:
♀ — родитель женского пола; ♂ — родитель мужского
пола; × - знак скрещивания;
P — родители; G (Г) — гаметы;
F - потомство (F1 - гибриды первого поколения,
F2 – гибриды второго поколения);
А – аллель гена, кодирующий доминантный признак,
а – аллель, кодирующий рецессивный признак;
АА – генотип доминантной гомозиготной особи,
аа – генотип гомозиготной рецессивной особи;
Аа – генотип гетерозиготной особи.
Первый - закон единообразия гибридов первого
поколения (закон доминирования) — при скрещивании
двух гомозиготных организмов, отличающихся друг от
друга по одной паре альтернативных проявлений
признака, всё первое поколение гибридов (F1) окажется
единообразным
и
будет
нести
проявление
признака одного из родителей.
Все потомство получится гетерозиготным и будет
проявлять действие доминантного аллеля. Этот закон
основан на варианте взаимодействия между генами полном доминировании, когда один ген - доминантный,
полностью подавляет другой ген - рецессивный., как в
случае с окраской семян гороха.
9.
Получившиеся в первом поколении потомки F1 (гибриды первого поколения) способныобразовывать два типа гамет: А и а, каждая из которых с равной вероятностью может
слиться с любой другой. В результате во втором поколении F2 образуются следующие
генотипы: АА, Аа, Аа, аа (1АА:2Аа:1aa). Рецессивный признак будет проявляться только у
особей аа, в то время как особи AA и Аа будут иметь доминантный признак. Таким
образом, расщепление (соотношение разных типов особей в потомстве) по генотипу
1:2:1, по фенотипу 3: 1.
Второй — закон расщепления — при скрещивании двух
гетерозиготных потомков первого поколения между
собой, во втором поколении наблюдается расщепление
в определенном числовом отношении: по фенотипу 3:1,
по генотипу 1:2:1.
При скрещивании организмов, отличающихся по двум парам аллелей A и а, B и b
(дигибридное скрещивание) образуется 4 типа гамет, в случае, если гены, кодирующие
2 признака, находятся в разных хромосомах, как в примере с окраской и формой семян
гороха. Это происходит из-за случайного расхождения гомологичных хромосом в
процессе мейоза (деления половых клеток). Расщепление по фенотипу 9:3:3:1
Третий — закон независимого распределения признаков — каждый ген одной
аллельной пары может оказаться в гамете с любым другим геном из другой аллельной
пары.
10.
СЦЕПЛЕНИЕ ГЕНОВТретий закон независимого наследования генов справедлив Опыт №1
лишь для генов, расположенных в разных хромосомах. На самом
деле число генов у эукариотов очень велико (десятки тысяч), а
число хромосом – ограничено, например, у человека -23 пары, у
кукурузы – 10, у мушки дрозофилы – 4 пары. Американский
ученый Томас Морган (1866-1945) проводил опыты по
дигибридному скрещиванию на дрозофиле, анализировал два
признака цвет тела и длину крыльев. Поскольку оба гена
расположены в одной хромосоме, образовывалось только 2 типа
гамет и расщепление было 3:1, а не 9:3:3:1, как ожидалось. При
анализирующем скрещивании (скрещивание с рецессивной
гомозиготой), появились особи со смешанными признаками.
Закон Томаса Моргана - гены, расположенные в одной хромосоме близко друг к другу,
образуют группу сцепления и наследуются совместно. Число сцепленных групп
соответствует гаплоидному набору – половине полного набора хромосом. Частота
наследования зависит от расстояния между генами. Чем ближе находятся гены,
образующие группы, тем чаще наследуются сцепленные признаки.
Гены одной хромосомы не обязательно наследуются вместе. Это связано с явлением
кроссинговера – обмена гомологичными хромосомами отдельными участками в
процессе мейоза. Аллели гомологичных хромосом меняются местами друг с другом.
Чем дальше на хромосоме расположены гены, тем чаще между ними происходит
обмен.
11.
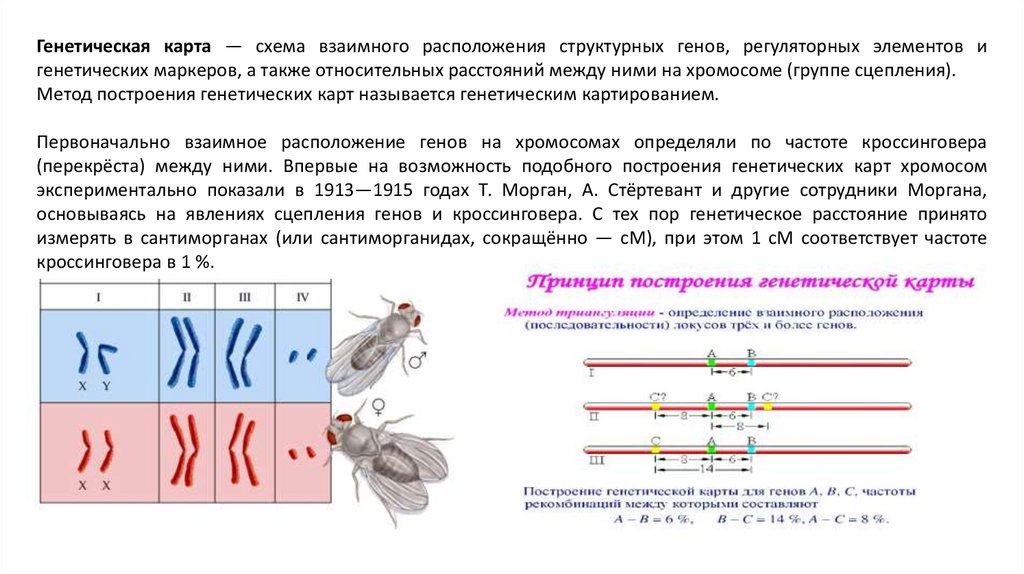
Генетическая карта — схема взаимного расположения структурных генов, регуляторных элементов игенетических маркеров, а также относительных расстояний между ними на хромосоме (группе сцепления).
Метод построения генетических карт называется генетическим картированием.
Первоначально взаимное расположение генов на хромосомах определяли по частоте кроссинговера
(перекрёста) между ними. Впервые на возможность подобного построения генетических карт хромосом
экспериментально показали в 1913—1915 годах Т. Морган, А. Стёртевант и другие сотрудники Моргана,
основываясь на явлениях сцепления генов и кроссинговера. С тех пор генетическое расстояние принято
измерять в сантиморганах (или сантиморганидах, сокращённо — cM), при этом 1 cM соответствует частоте
кроссинговера в 1 %.
12.
13.
Цитогенетическая карта — пространственное представлениепорядка взаимного расположения структурных элементов хромосом
(например,
локусов
гибридизации
меченых
ДНК-зондов
(флуоресцентная гибридизация in situ).
Физическая карта — представление порядка следования физических
маркеров (фрагментов молекулы ДНК), расстояние между которыми
определяется в парах нуклеотидов (п. н.).
Рестрикционная карта — вид физической карты, на которой указан
порядок следования и расстояния между сайтами расщепления ДНКрестриктазами (обычно участок узнавания рестриктазы размером
4—6 п. н.). Маркерами этой карты являются рестрикционные
фрагменты (сайты рестрикции).
Конечной целью изучения генома данного организма является
интеграция его генетических, цитогенетических и физических карт, а
также их привязка к полной геномной последовательности.
14.
15.
В 1869 г. И. Мишером была открыта ДНК. Биологическая функция нового вещества была не ясна.В 1944 г. ученые О. Эвери, К. Маклеод и М. Маккарти провели ряд экспериментов по трансформации
бактерий и доказали, что за трансформацию (приобретение болезнетворных свойств безвредной культурой в
результате добавления в нее мертвых болезнетворных бактерий) отвечают выделенные из пневмококков
ДНК.
1949-1951 – правило Чаргаффа – количественные соотношения между различными типами азотистых
оснований в составе нуклеотидов ДНК. Количество пуринов равно количеству пиримидинов: А + Г = Т + Ц.
1953 г. Дж.Уотсон, Ф.Крик – открытие структуры ДНК.
16.
В 1955 г. А. Корнберг открыл фермент, который назвал ДНК-полимеразой. Этот фермент способен удлинятьцепь ДНК, присоединяя нуклеотиды к ее 3'-концу. В искусственных условиях фермент катализирует реакцию
достраивания участка искомой ДНК от затравки (праймера), которая комплементарно связана с цепью ДНК
(матрицей). Раствор, в котором происходит эта реакция, должен содержать нуклеозидтрифосфаты (дНТФ),
используемые в качестве строительных блоков.
1958 г. Фрэнсис Крик сформулировал так называемую центральную догму молекулярной биологии:
представление о необратимости потока генетической информации от ДНК через РНК к белкам по схеме
ДНК→ДНК (репликация, создание копии ДНК), ДНК→РНК (транскрипция), РНК→ белок (трансляция,
декодирование информации о структуре белков).
1970 г. открыто явление обратной транскрипции
независимо Ховардом Темином и Дэвидом
Балтимором: был обнаружен фермент –
ревертаза, отвечающий за осуществление
обратной
транскрипции
–
образования
двуцепочечной
ДНК
на
матрице
одноцепочечной РНК, которое происходит у
онкогенных вирусов.
17.
В 1975 г. Т. Брок и Х. Фриз открыли Thermus aquaticus – грамотрицательную палочковидную экстремальнотермофильную бактерию, а в 1976 г. из нее была впервые выделена Taq-полимераза. Преимуществом
данного фермента была способность стабильно работать при повышенных температурах (оптимум 72-80 °C).
В 1983-1984 гг. К. Мюллис провел ряд экспериментов по разработке ПЦР и первым начал использовать Taqполимеразу вместо неустойчивой к высоким температурам ДНК-полимеразы. Это позволило ускорить работы
по разработке полимеразной цепной реакции. Кроме того, К. Мюллис вместе с Ф. Фалуном разработали
алгоритм циклических изменений температуры в ходе ПЦР.
18.
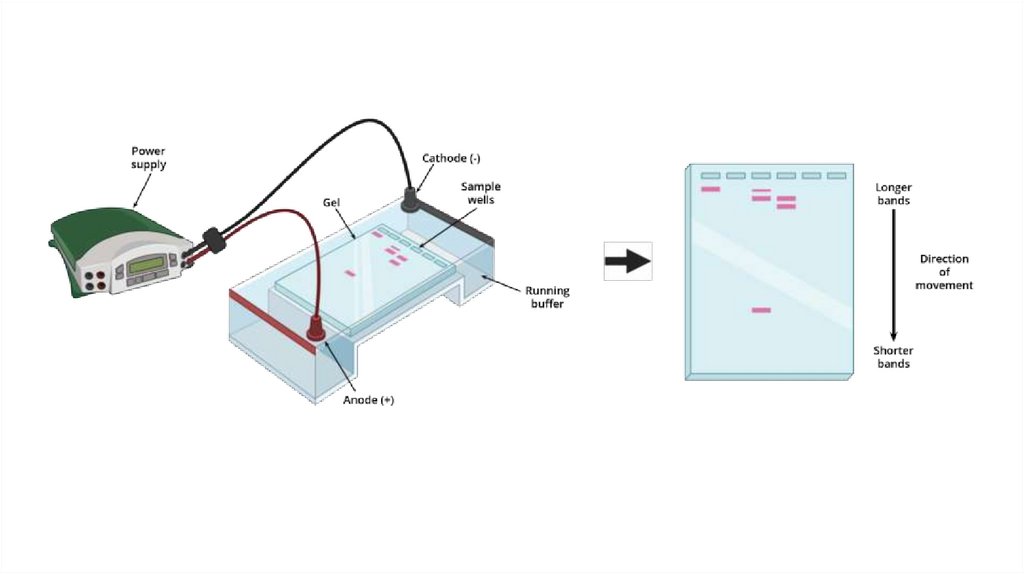
Полимеразная цепная реакция (ПЦР) – экспериментальный метод молекулярнойбиологии, позволяющий добиться значительного увеличения малых концентраций
определенных фрагментов нуклеиновой кислоты (НК) в биологическом материале
(пробе).
19.
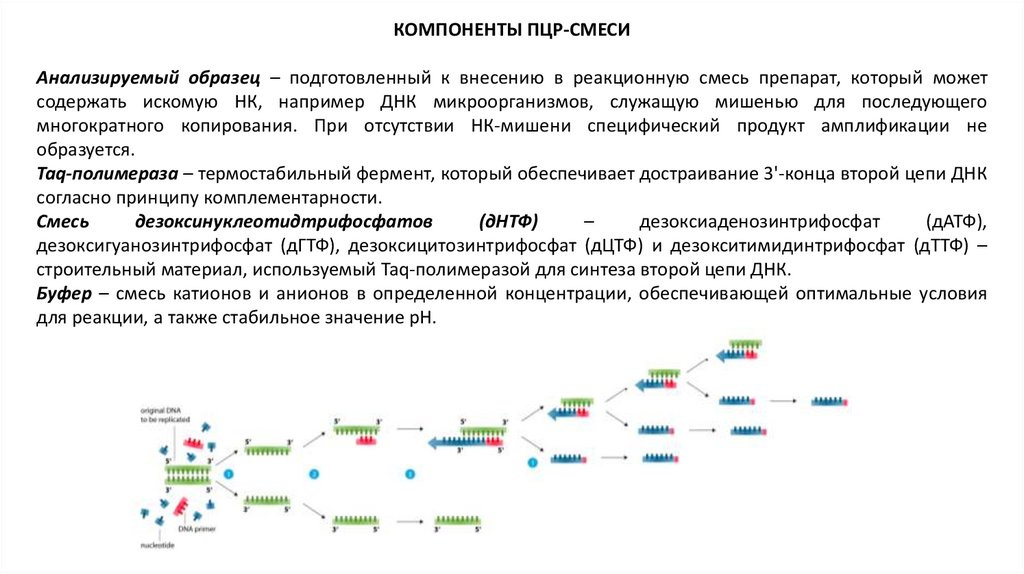
КОМПОНЕНТЫ ПЦР-СМЕСИАнализируемый образец – подготовленный к внесению в реакционную смесь препарат, который может
содержать искомую НК, например ДНК микроорганизмов, служащую мишенью для последующего
многократного копирования. При отсутствии НК-мишени специфический продукт амплификации не
образуется.
Taq-полимераза – термостабильный фермент, который обеспечивает достраивание 3'-конца второй цепи ДНК
согласно принципу комплементарности.
Смесь
дезоксинуклеотидтрифосфатов
(дНТФ)
–
дезоксиаденозинтрифосфат
(дАТФ),
дезоксигуанозинтрифосфат (дГТФ), дезоксицитозинтрифосфат (дЦТФ) и дезокситимидинтрифосфат (дТТФ) –
строительный материал, используемый Taq-полимеразой для синтеза второй цепи ДНК.
Буфер – смесь катионов и анионов в определенной концентрации, обеспечивающей оптимальные условия
для реакции, а также стабильное значение рН.
20.
Праймеры – искусственно синтезированные олигонуклеотиды, имеющие, как правило, размер от 15 до 30нуклеотидов, идентичные (комплементарные) противоположным концам противоположных цепей искомого
участка ДНК-мишени. Служат затравкой для синтеза комплементарной цепи с помощью ДНК-полимеразы и
играют ключевую роль в образовании и накоплении продуктов реакции амплификации.
Требования к праймерам:
должны быть специфичными;
не должны образовывать димеры и петли – устойчивые двойные цепи - при отжиге праймеров самих на
себя или друг с другом;
область отжига праймеров должна находиться вне зон мутаций, делеций или инсерций в пределах
видовой или иной специфичности, взятой в качестве критерия при выборе праймеров. При попадании в
такую зону не происходит отжига праймеров и, как следствие, возникает ложноотрицательный результат.
21.
1. Денатурация – это переход ДНК из двухнитевойформы в однонитевую при
разрыве
водородных
связей
между
комплементарными
парами
оснований
противоположных цепей ДНК под воздействием
высоких температур.
2. Отжиг – это присоединение праймеров к
одноцепочечной ДНК-мишени.
Праймеры подбирают так, чтобы они ограничивали
искомый фрагмент ДНК и были комплементарны
противоположным цепям ДНК. Отжиг происходит в
соответствии с правилом комплементарности
Чаргаффа. Если это условие не соблюдено, то отжига
праймеров не происходит.
3. Элонгация (синтез). После отжига праймеров Taqполимераза начинает достраивание второй цепи ДНК
с 3'-конца праймера с использованием дНТФ.
22.
ОТ-ПЦР широко используется для выявления вирусов, геном которых представлен РНК (ВИЧ, гепатит С, вирусыгриппа и другие), для диагностики генетических заболеваний и полуколичественного определения
специфических молекул РНК в клетке или ткани как индикатор экспрессии соответствующих генов.