








































 biology
biologySimilar presentations:








")
")
Физиология мышц. Структурно-функциональная характеристика скелетной мышцы и механизм ее сокращения
1. ФИЗИОЛОГИЯ МЫШЦ
СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯХАРАКТЕРИСТИКА СКЕЛЕТНОЙ
МЫШЦЫ И МЕХАНИЗМ ЕЕ
СОКРАЩЕНИЯ
2.
Структурной единицей скелетной мышцыявляется мышечное волокно - сильно вытянутая
многоядерная клетка.
Длина мышечного волокна зависит от размеров
мышцы и составляет от нескольких миллиметров
до нескольких сантиметров. Толщина волокна
варьирует от (10-100 мкм).
Типы мышц
В организме человека существует три типа
мышц:
скелетные, сердечные (миокард) и гладкие.
При микроскопическом исследовании в
скелетных и сердечной мышцах
обнаруживается исчерченностъ, поэтому их
называют поперечнополосатыми мышцами.
3.
Скелетные мышцы прикреплены в основном ккостям, что и обусловило их название.
Сокращение скелетных мышц инициируется
нервными
импульсами
и
подчиняется
сознательному
контролю,
т.е.
осуществляется произвольно.
Сокращение гладких мышц инициируется
импульсами, некоторыми гормонами и не
зависит от воли человека.
4.
Мышечное волокно окружено двухслойнойлипопротеидной электровозбудимой мембраной сарколеммой,
которая
покрыта
сетью
коллагеновых волокон, придающих ей прочность и
эластичность.
В скелетных мышцах различают несколько типов
мышечных волокон: медленносокращающиеся
(МС) или красные и быстросокращающиеся
(БС) или белые.
Молекулярный механизм сокращения.
Скелетные мышцы содержат сократительные
белки:
актин
и
миозин.
Механизм
их
взаимодействия во время элементарного акта
мышечного
сокращения
объясняет
теория
скользящих нитей, разработанная Хасли и
Хансоном.
5. Строение мышечного волокна
Сарколемма – плазматическая мембрана покрывающаямышечное волокно (соединяется с сухожилием, которое
прикрепляет мышцу к кости; сухожилие передает усилие
производимое мышечными волокнами кости и таким
образом
осуществляется
движение).
Сарколемма
обладает избирательной проницаемостью для различных
веществ и имеет транспортные системы, с помощью
которых поддерживается разная концентрация ионов
Na+, К+, а также Сl- внутри клетки и в межклеточной
жидкости, что приводит к возникновению на ее
поверхности мембранного потенциала - необходимого
условия возникновения возбуждения мышечного волокна.
Саркоплазма
–
желатиноподобная
жидкость,
заполняющая
промежутки
между
миофибриллами
(содержит
растворенные
белки,
микроэлементы,
гликоген, миоглобин, жиры, органеллы). Около 80%
объема волокна занимают длинные сократительные нити
- миофибриллы.
6.
Система поперечных трубочек. Это сеть Т –трубочек (поперечные), является продолжением
сарколеммы; они взаимосоединяются проходя
среди миофибрилл. Обеспечивают быструю
передачу нервных импульсов (распространение
возбуждения) внутрь клетки к отдельным
миофибриллам.
Саркоплазматический ретикулум (СР) – сеть
продольных трубочек, расположены параллельно
миофибриллам; это место депонирования Са2+,
который необходим для обеспечения процесса
мышечного сокращения.
Сократительные белки актин и миозин образуют
в миофибриллах тонкие и
толстые
миофиламенты.
Они
располагаются
параллельно друг другу внутри мышечной клетки
Миофибриллы
представляют
собой
сократимые элементы мышечного волокна пучки «нитей» (филаментов).
7.
Структура миофибриллы:1. Перегородки – называемые Z - пластинками,
разделяют их на саркомеры.
Структура саркомера:
В них видна последовательность регулярно
чередующихся поперечных светлых и темных
полос,
которая
обусловлена
особым
взаиморасположением
актиновых
и
миозиновых
филаментов
(поперечная
полосатость).
Середину саркомера занимают «толстые» нити
миозина. ( А – диск темный)
На
обоих концах саркомера находятся
«тонкие» нити актина. (I- диск светлый)
8.
Актиновые нити прикрепляются к Z –пластинкам, сами Z – пластинки
ограничивают саркомер.
В покоящейся мышце концы тонких и
толстых
филаментов
лишь
слабо
перекрываются на границе между А и Iдисками.
Н – зона (светлее) в которой нет
перекрывания
нитей
(здесь
располагаются только миозиновые нити),
находится в диске А.
М - линия находится в центре саркомера
– место удержания толстых нитей
(построена из опорных белков.)
9. Теория скользящих нитей.
Укорочение саркомера:Мышца сокращается в результате укорочения множества
последовательно соединенных саркомеров в
миофибриллах.
Во время сокращения тонкие актиновые филаменты
скользят вдоль толстых миозиновых, двигаясь между ними
к середине их пучка и саркомера.
Основное положение теории скользящих нитей:
Во время сокращения мышцы, сами актиновые и
миозиновые нити не укорачиваются (ширина А – диска
всегда остается постоянной, тогда как I- диски и Н – зоны
при сокращении сужаются).
Длинна нитей не меняется при растяжении мышцы (тонкие
филаменты вытягиваются из промежутков между толстыми
нитями, так что степень перекрывания их пучков
уменьшается).
10. Работа поперечных мостиков.
Движение головок создает объединенное усилие,как бы «гребок», продвигающий актиновые нити к
середине саркомера. Только за счет ритмичных
отделений и повторных прикреплений миозиновых
головок актиновая нить может подтягиваться к
середине саркомера.
При расслаблении мышцы миозиновые головки
отделяются от актиновых нитей.
Так как актиновые и миозиновые нити могут легко
скользить друг относительно друга, сопротивление
расслабленных мышц растяжению очень низкое.
Удлинение мышцы во время расслабления носит
пассивный характер.
11. Преобразование химической энергии в механическую.
АТФ – непосредственный источник энергии длясокращения.
При сокращении мышцы АТФ расщепляется на
АДФ и фосфат.
Ритмическая активность поперечных мостиков, т.
е. циклы их прикрепления к актину и отсоединения
от него , обеспечивающие мышечное сокращение,
возможны только при гидролизе АТФ , а
соответственно и при активации АТФазы, которая
непосредственно участвует в расщеплении АТФ на
АДФ и фосфат.
12. Молекулярный механизм мышечного сокращения.
Сокращение запускается нервным импульсом. При этом всинапсе - месте контакта нервного окончания с
сарколеммой выделяется медиатор (нейропередатчик) ацетилхолин.
Ацетилхолин (Ах) вызывает изменение проницаемости
мембраны для некоторых ионов, что в свою очередь
приводит к возникновению ионных токов и сопровождается
деполяризацией мембраны. В следствии чего , на ее
поверхности возникает потенциал действия или она
возбуждается.
Потенциал
действия
(возбуждение)
распространяется вглубь волокна через Т-системы.
Нервный импульс вызывает изменение проницаемости
мембраны саркоплазматического ретикулума и приводит к
освобождению
ионов
Са2+
из
пузырьков
саркоплазматического ретикулума.
13. Электромеханическое сопряжение
Передача команды к сокращению отвозбужденной клеточной мембраны к
миофибриллам
в
глубине
клетки
(электромеханическое
сопряжение)
включает
в
себя
несколько
последовательных процессов, ключевую
роль в которых играют ионы Са2+.
14.
1. Электромеханическое сопряжение происходитпосредством распространения потенциала
действия по мембранам поперечной системы
внутрь клетки, потом возбуждение проходит на
продольную систему (ЭПР ) и вызывает
высвобождение депонированного в мышечной
клетке Са2+ во внутриклеточное пространство,
которое окружает миофибриллы. Это и приводит к
сокращению
2. Са2+ удаляется из внутриклеточного пространства
в депо (каналы ЭПР) за счет работы кальциевых
насосов на мембранах ЭПР.
3. Только за счет электрической передачи по
поперечной системе, возможна быстрая
мобилизация запасов кальция в глубине волокна, и
только этим можно объяснить очень короткий
латентный период между стимулом и
сокращением.
15.
Функциональная роль АТФ :- в покоящейся мышце - препятствует соединению
актиновых нитей с миозиновыми;
- в процессе сокращения мышцы - поставляет
необходимую энергию для движения тонких нитей
относительно толстых, что приводит к укорочению
мышцы или развитию напряжения;
- в процессе расслабления - обеспечивает энергией
активный транспорт Са2+ в ретикулум.
16. Типы мышечных сокращений. Оптимум и пессимум мышечного сокращения
В зависимости от изменения длины мышечного волокнавыделяют два типа его сокращения - изометрическое и
изотоническое.
Мышечное сокращение при котором длина мышцы
уменьшается по мере развиваемой ею силы, называется
ауксотоническим.
Максимальная сила при ауксотонических экспериментальных
условиях (с растяжимой упругой связью между мышцей и
датчиком силы) называется максимумом ауксотонического
сокращения. Она гораздо меньше силы, которую развивает
мышца при постоянной длине, т.е. при изометрическом
сокращении.
Сокращение мышцы, при котором ее волокна укорачиваются
при неизменном напряжении, называется изотоническим.
Сокращение мышцы, при котором ее напряжение возрастает
а длина мышечных волокон остается неизменной,
называется изометрическим
17.
Мышечная работа равна произведениюрасстояния (укорочения мышцы) на вес груза,
который поднимает мышца.
При изотонической тетанической активации
мышцы от нагрузки зависит величина укорочения и
скорость укорочения мышцы.
Чем меньше нагрузка, тем больше укорочений в
единицу времени. Ненагруженная мышца
укорачивается с максимальной скоростью, которая
зависит от типа мышечных волокон.
Мощность мышцы равна произведению
развиваемой ею силы на скорость укорочения
18.
Расслабленная мышца, сохраняющая «длину покоя» за счетфиксации обоих ее концов, не развивает силу, которая
передавалась бы на датчик. Но если потянуть за один ее
конец, чтобы волокна растянулись, в ней возникает
пассивное напряжение. Таким образом, мышца в состоянии
покоя упруга. Модуль упругости покоящейся мышцы с
растяжением возрастает. Эта упругость обусловлена главным
образом растяжимыми структурами, которые располагаются
параллельно
относительно
растяжимых
миофибрилл
(«параллельная
упругость»)
.
Миофибриллы
в
расслабленном состоянии практически не оказывают
сопротивления растяжению; актиновые и миозиновые нити, не
связанные
поперечными
мостиками,
легко
скользят
относительно друг друга. Степень предварительного
растяжения определяет величину пассивного напряжения
покоящейся мышцы и величину дополнительной силы,
которую может развить мышца в случае активации при данной
длине.
19.
Пиковое усилие при таких условиях называетсямаксимумом изометрического сокращения.
При сильном растяжении мышцы, сила сокращения
уменьшается т. к. нити актина вытянуты из
миозиновых пучков и соответственно, меньше зона
перекрывания этих нитей и возможность
формирования поперечных мостиков.
При очень сильном растяжении мышцы, когда
актиновые и миозиновые нити перестают
перекрываться , миофибриллы не способны
развивать силу. Это доказывает, что мышечная сила
представляет собой результат взаимодействия
актиновых и миозиновых филаментов ( т. е.
образования между ними поперечных мостиков).
В естественных условиях сокращения мышц
являются смешанными - мышца обычно не только
укорачивается, но изменяется и ее напряжение.
20.
В зависимости от длительности выделяютодиночное и тетаническое сокращения мышцы.
Одиночное сокращение мышцы в эксперименте
вызывают одиночным раздражением электрическим
током. В изотоническом режиме одиночное
сокращение начинается через короткий скрытый
(латентный) период, далее следует фаза подъема
(фаза укорочения), затем фаза спада (фаза
расслабления) (рис. 1). Обычно мышца
укорачивается на 5-10% исходной длины.
Длительность ПД мышечных волокон также
варьирует и составляет 5-10 мс с учетом замедления
фазы реполяризации.
Мышечное волокно подчиняется закону «все или
ничего», т.е. отвечает на пороговое и
сверхпороговое раздражение одинаковым по
величине одиночным сокращением.
21.
Сокращение целой мышцы зависит:1. от силы раздражителя при непосредственном раздражении
мышцы
2. от числа нервных импульсов, поступающих к мышце при
раздражении нерва.
Увеличение силы раздражителя ведет к увеличению числа
сокращающихся мышечных волокон.
Подобный эффект наблюдается и в естественных условиях - с
увеличением числа возбужденных нервных волокон и частоты
импульсов (к мышце поступает больше нервных импульсов ПД) увеличивается число сокращающихся мышечных волокон.
При одиночных сокращениях мышца утомляется
незначительно.
Тетаническое сокращение - это слитное длительное
сокращение скелетной мышцы. В его основе лежит явление
суммации одиночных мышечных сокращений.
Кривая одиночного
сокращения икроножной
мышцы лягушки:
1-латентный период,
2- фаза укорочения,
22.
При нанесении на мышечное волокно илинепосредственно
на
мышцу
двух
быстро
следующих друг за другом раздражений ,
возникающее
сокращение
имеет
большую
амплитуду и длительность. При этом нити актина и
миозина дополнительно скользят друг относительно
друга. В сокращение могут вовлекаться ранее не
сокращавшиеся мышечные волокна, если первый
стимул вызвал у них подпороговую деполяризацию,
а второй увеличивает ее до критической величины.
Суммация сокращений при повторном раздражении
мышцы или поступлении к ней ПД возникает только
в том случае, когда закончен рефрактерный период
(после исчезновения ПД мышечного волокна).
23.
При поступлении импульсов к мышце во время еерасслабления возникает зубчатый тетанус, во
время укорочения - гладкий тетанус (рис.).
Амплитуда тетануса больше величины
максимального одиночного сокращения мышцы.
Напряжение, развиваемое мышечными волокнами
при гладком тетанусе, обычно в 2-4 раза больше,
чем при одиночном сокращении, однако мышца
быстрее утомляется. Мышечные волокна не
успевают восстановить энергетические ресурсы,
израсходованные во время сокращения.
Амплитуда гладкого тетануса увеличивается с
возрастанием частоты стимуляции нерва. При
некоторой (оптимальной) частоте стимуляции
амплитуда гладкого тетануса наибольшая (оптимум частоты раздражения)
24.
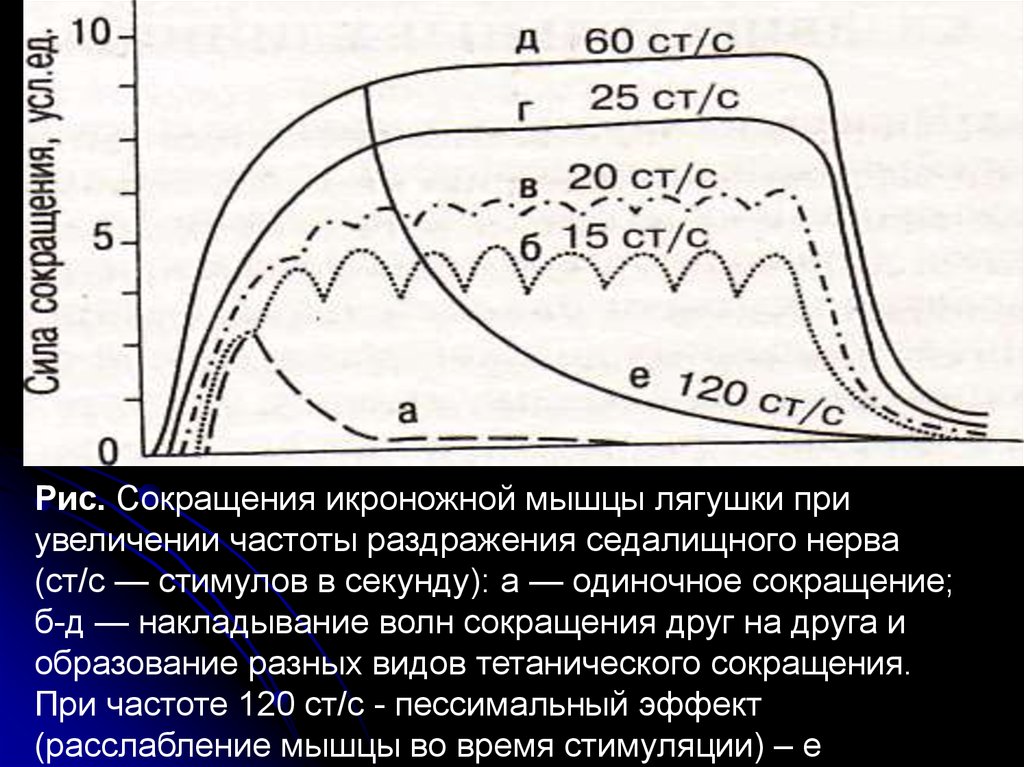
Рис. Сокращения икроножной мышцы лягушки приувеличении частоты раздражения седалищного нерва
(ст/с — стимулов в секунду): а — одиночное сокращение;
б-д — накладывание волн сокращения друг на друга и
образование разных видов тетанического сокращения.
При частоте 120 ст/с - пессимальный эффект
(расслабление мышцы во время стимуляции) – е
25.
При чрезмерно частой стимуляции нерва (более 100имп/с) мышца расслабляется вследствие блока
проведения возбуждения в нервно-мышечных
синапсах - пессимум Введенского (пессимум
частоты раздражения). Пессимум Введенского можно
получить и при прямом, но более частом раздражении
мышцы (более 200 имп/с) . Пессимум Введенского не
является результатом утомления мышцы или истощения медиатора в синапсе, что доказывается фактом
возобновления сокращения мышцы сразу же после
уменьшения частоты раздражения. Торможение
развивается в нервно-мышечном синапсе при
раздражении нерва.
В естественных условиях мышечные волокна
сокращаются в режиме зубчатого тетануса или
даже одиночных последовательных сокращений.
26.
Однако форма сокращения мышцы в целомнапоминает гладкий тетанус.
Причины
этого
асинхронность
разрядов
мотонейронов и асинхронность сократительной
реакции отдельных мышечных волокон, вовлечение
в сокращение большого их количества, вследствие
чего мышца плавно сокращается и плавно
расслабляется, может длительно находиться в
сокращенном состоянии за счет чередования
сокращений множества мышечных волокон. При
этом мышечные волокна каждой двигательной
единицы сокращаются синхронно.
27.
Функциональная единица мышцы –двигательная единица
Понятия. Иннервация скелетных мышечных волокон
осуществляется мотонейронами спинного мозга или
мозгового ствола. Один мотонейрон веточками своего
аксона иннервирует несколько мышечных волокон.
Совокупность мотонейрона и иннервируемых им
мышечных волокон называют двигательной
(нейромоторной) единицей. Число мышечных
волокон двигательной единицы варьирует в широких
пределах в разных мышцах. Двигательные единицы
невелики в мышцах, приспособленных для быстрых
движений, от нескольких мышечных волокон до
нескольких десятков их (мышцы пальцев, глаза,
языка). Наоборот, в мышцах, осуществляющих
медленные движения (поддержание позы мышцами
туловища), двигательные единицы велики и включают
сотни и тысячи мышечных волокон
28.
Присокращении
мышцы
в
натуральных
(естественных) условиях можно зарегистрировать
ее электрическую активность (электромиограмму ЭМГ) с помощью игольчатых или накожных электродов. В абсолютно расслабленной мышце
электрическая активность почти отсутствует. При
небольшом
напряжении,
например
при
поддержании
позы,
двигательные
единицы
разряжаются с небольшой частотой (5-10 имп/с),
при большом напряжении частота импульсации
повышается в среднем до 20-30 имп/с. ЭМГ позволяет судить о функциональной способности
нейромоторных единиц. С функциональной точки
зрения двигательные единицы разделяют на
медленные и быстрые.
29.
мотонейроны и медленные мышечные волокна (красные).Медленные мотонейроны, как правило, низкопороговые, так
как обычно это малые мотонейроны. Устойчивый уровень
импульсации у медленных мотонейронов наблюдается уже
при очень слабых статических сокращениях мышц, при
поддержании позы. Медленные мотонейроны способны
поддерживать длительный разряд без заметного снижения
частоты импульсации на протяжении длительного времени.
Поэтому их называют малоутомляемыми или
неутомляемыми мотонейронами. В окружении медленных
мышечных волокон богатая капиллярная сеть, позволяющая
получать большое количество кислорода из крови.
Повышенное содержание миоглобина облегчает транспорт
кислорода в мышечных клетках к митохондриям. Миоглобин
обусловливает красный цвет этих волокон. Кроме того,
волокна содержат большое количество митохондрий и
субстратов окисления - жиров. Все это обусловливает использование медленными мышечными волокнами более
эффективного аэробного окислительного пути
30.
Быстрые двигательные единицы состоят избыстрых мотонейронов и быстрых мышечных
волокон. Быстрые высокопороговые мотонейроны
включаются в активность только для обеспечения
относительно больших по силе статических и
динамических сокращений мышц, а также в начале
любых сокращений, чтобы увеличить скорость
нарастания напряжения мышцы или сообщить
движущейся части тела необходимое ускорение. Чем
больше скорость и сила движений, т. е. чем больше
мощность сократительного акта, тем больше участие
быстрых двигательных единиц. Быстрые
мотонейроны относятся к утомляемым - они не
способны к длительному поддержанию
высокочастотного разряда
31.
Быстрые мышечные волокна (белые мышечныеволокна) более толстые, содержат больше
миофибрилл, обладают большей силой, чем
медленные волокна. Эти волокна окружает меньше
капилляров, в клетках меньше митохондрий,
миоглобина и жиров. Активность окислительных
ферментов в быстрых волокнах ниже, чем в
медленных, однако активность гликолитических
ферментов, запасы гликогена выше. Эти волокна не
обладают большой выносливостью и более
приспособлены для мощных, но относительно
кратковременных сокращений. Активность быстрых
волокон имеет значение для выполнения
кратковременной высокоинтенсивной работы,
например бега на короткие дистанции
32.
Скорость сокращения мышечных волокон находитсяв прямой зависимости от активности миозин-АТФ-азы
- фермента, расщепляющего АТФ и тем самым
способствующего образованию поперечных мостиков
и взаимодействию актиновых и миозиновых
миофиламентов. Более высокая активность этого
фермента в быстрых мышечных волокнах
обеспечивает и более высокую скорость их
сокращения по сравнению с медленными волокнами
Тонус – слабое общее напряжение мышц
(развивается при очень низкой частоте стимуляции).
Сила и скорость сокращения мышц зависит от
количества вовлеченных в сокращение двигательных
единиц (чем больше двигательных единиц
активировано – тем сильнее сокращение).
Рефлекторный тонус - (наблюдается у некоторых
групп позных мышц) состояние непроизвольного
устойчивого напряжения мышц
33.
КПД мышцыВо время активации мышцы повышение
внутриклеточной концентрации Са 2+ ведет к
сокращению и к усиленному расщеплению АТФ; при
этом интенсивность метаболизма мышцы возрастает
в 100-1000 раз. Согласно первому началу
термодинамики (закону сохранения энергии),
химическая энергия, высвобождаемая в мышце,
должна быть равна сумме механической энергии
(мышечной работы) и теплообразования
34.
Коэффициент полезного действия.Гидролиз одного моля АТФ дает 48 кДж энергии,
40 –50% - превращается в механическую работу, а
50-60% рассеивается в виде тепла при запуске
(начальная теплота) и во время сокращения
мышцы, температура которой при этом
повышается. Однако в естественных условиях
механический КПД мышц около 20-30% так как во
время сокращения и после него процессы
требующие затрат энергии, идут и вне
миофибрилл (работа ионных насосов,
окислительная регенерация АТФ – теплота
восстановления)
35.
Энергетическийметаболизм
.
Во
время
продолжительной
равномерной
мышечной
активности происходит аэробная регенерация АТФ за
счет
окислительного
фосфорильирования.
Необходимая для этого энергия выделяется в
результате окисления углеводов и жиров. Система
находится в состоянии динамического равновесия –
скорости образования и расщепления АТФ равны.
(внутриклеточные
концентрации
АТФ
и
креатинфосфата относительно постоянны) При
продолжительных спортивных нагрузках скорость
расщепления АТФ в мышцах возрастает в 100 или в
1000 раз. Продолжительная нагрузка возможна если
скорость
восстановления
АТФ
возрастает
соответственно расходу. Потребление кислорода
мышечной тканью возрастает в 50-100 раз;
повышается скорость расщепления гликогена в
мышцах.
36.
Анаэробное расщепление – гликолиз: АТФ образуется в 2-3раза быстрее, а механическая энергия мышцы в 2-3 раза
выше, чем при длительной работе , обеспечиваемой
аэробными механизмами. Но ресурсы для анаэробного
метаболизма быстро исчерпываются, продукты метаболизма
(молочная кислота) вызывают метаболический ацидоз.,
который ограничивает работоспособность и вызывает
утомление. Анаэробные процессы необходимы для
обеспечения энергией кратковременного экстремального
усилия, а так же в начале продолжительной мышечной
работы, потому что адаптация скорости окисления ( и
гликолиза) к возросшей нагрузке требует некоторого времени.
Кислородная задолженность приблизительно соответствует
количеству энергии, полученному анаэробным путем, еще не
компенсированное за счет аэробного синтеза АТФ.
Кислородная задолженность обусловлена(анаэробным )
гидролизом креатинфосфата, может достигать 4 л и может
увеличиваться до 20 л. Часть лактата окисляется в миокарде
а часть(преимущественно в печени) используется для синтеза
гликогена.
37.
Соотношение быстрых, и медленных волокон. Чембольше быстрых волокон содержит мышца, тем больше
возможная ее сила сокращения.
Поперечное сечение мышцы.
Термины «абсолютная» и «относительная» сила мышцы :
«общая сила мышцы» (определяется максимальным
напряжением в кг, которое она может развить) и «удельная
сила мышцы» - отношение этого напряжения в кг к
физиологическому поперечному сечению мышцы (кг/см2).
Чем больше физиологическое поперечное сечение мышцы,
тем больший груз она в состоянии поднять. По этой причине
сила мышцы с косо расположенными волокнами больше
силы, развиваемой мышцей той же толщины, но с
продольным расположением волокон. Для сравнения силы
разных мышц максимальный груз, который они в состоянии
поднять, делят на плошадь их физиологического поперечного
сечения (удельная сила мышцы). Вычисленная таким образом
сила (кг/см2) для трехглавой мышцы плеча человека - 16,8,
двуглавой мышцы плеча - 11,4, сгибателя плеча — 8,1,
икроножной мышцы - 5,9, гладких мышц - 1 кг/см2.
38.
В различных мышцах тела соотношение междучислом медленных и быстрых мышечных волокон
неодинаково, поэтому и сила их сокращения, и
степень укорочения вариабельны.
При снижении физической нагрузки - особенно
большой интенсивности, при которой требуется
активное участие быстрых мышечных волокон, последние истончаются (гипотрофируются) быстрее,
чем медленные волокна, быстрее уменьшается их
число
Факторы, влияющие на силу сокращения мышцы.
Число сокращающихся волокон в данной мышце. С
увеличением сокращающихся волокон возрастает
сила сокращений мышцы в целом. В естественных
условиях сила сокращения мышцы возрастает с
увеличением нервных импульсов, поступающих к
мышце,
в эксперименте - с увеличением силы раздражения.
39.
Умеренное растяжение мышцы также ведет кувеличению ее сократительного эффекта. Однако
при чрезмерном растяжении сила сокращения
уменьшается. Это демонстрируется в опыте с
дозированным растяжением мышцы: мышца
перерастянута так, что нити актина и миозина не
перекрываются, то общая сила мышцы равна нулю.
По мере приближения к натуральной длине покоя,
при которой все головки миозиновых нитей способны
контактировать с актиновыми нитями, сила
мышечного сокращения вырастает до максимума.
Однако при дальнейшем уменьшении длины
мышечных волокон из-за перекрытия нитей актина и
миозина сила сокращения мышцы снова
уменьшается вследствие уменьшения возможной
зоны контакта нитей актина и миозина.
40.
Функциональное состояние мышцы.При утомлении мышцы величина ее сокращения
снижается.
Работа мышцы измеряется произведением
поднятого груза на величину ее укорочения.
Зависимость мышечной работы от нагрузки
подчиняется закону средних нагрузок. Если мышца
сокращается без нагрузки, ее внешняя работа равна
нулю. По мере увеличения груза работа
увеличивается, достигая максимума при средних
нагрузках. Затем она постепенно уменьшается с
увеличением нагрузки. Работа становится равной
нулю при очень большом грузе, который мышца при
своем сокращении не способна поднять напряжение
100-200 мг.
41.
ГЛАДКАЯ МЫШЦА.Гладкая мускулатура не имеет поперечную
исчерченность. Клетки в виде веретен соединены
особыми межклеточными контактами (десмосомами).
Скорость скольжения миофибрилл и расщепления АТФ
ниже в 100-1000 раз. Хорошо приспособлены для
длительного устойчивого сокращения, которое не
приводит к утомлению и значительным энергозатратам.
Способны к спонтанным тетанообразным сокращениям,
которые имеют миогенное происхождение, а не
нейрогенное как у скелетных мышц.
Миогенное возбуждение.
Миогенное возбуждение возникает в клетках
ритмоводителях (пейсмекерах), которые обладают
электрофизиологическими свойствами.
Пейсмекерные потенциалы деполяризуют их мембрану
до порогового уровня, вызывая потенциал действия. Са
2+ поступает в клетку – мембрана деполяризуется, потом
42.
Спонтанную активность пейсмекеров можно модулироватьвегетативной нервной системой и ее медиаторами
(ацетилхолин усиливает активность приводя к более частым и
сильным сокращениям, а норадреналин оказывает
противоположное действие).
Возбуждение распространяется через «щелевые контакты»
(нексусы) между плазматическими мембранами
сопредельных мышечных клеток. Мышца ведет себя как
единая функциональная единица, синхронно воспроизводя
активность своего пейсмекера. Гладкая мышца может быть
полностью расслаблена как в укороченном так и в растянутом
состоянии. Сильное растяжение активирует сокращение.
Электромеханические сопряжение. Возбуждение
гладкомышечных клеток вызывает либо увеличение входа Са
через потенциалзависимые кальциевые каналы, либо
высвобождает из кальциевых депо, что в любом случае
приводит к возрастанию внутриклеточной концентрации
кальция и вызывает активацию сократительных структур.
Расслабление идет медленно т.к. скорость поглощения ионов
Са очень низкая.