




 biology
biologySimilar presentations:
")









Генетическая рекомбинация
1.
ГЕНЕТИЧЕСКАЯ РЕКОМБИНАЦИЯ2.
РЕКОМБИНАЦИЯ•Общая (гомологичная) – происходит между гомологичными последовательностями
ДНК, у эукариот в мейозе или реже митозе.
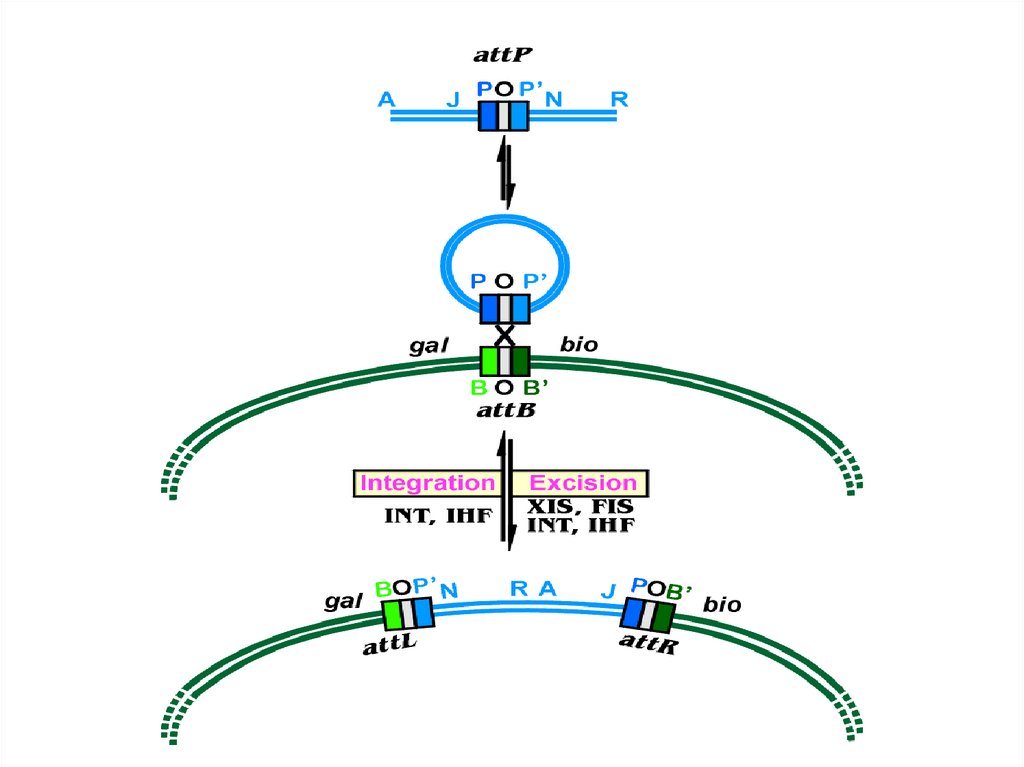
•Сайт-специфическая – затрагивает ограниченные последовательности ДНК со
структурным сходством
•Незаконная – не основана на гомологии последовательностей ДНК,
происходит во время транспозиции
Генная конверсия
3.
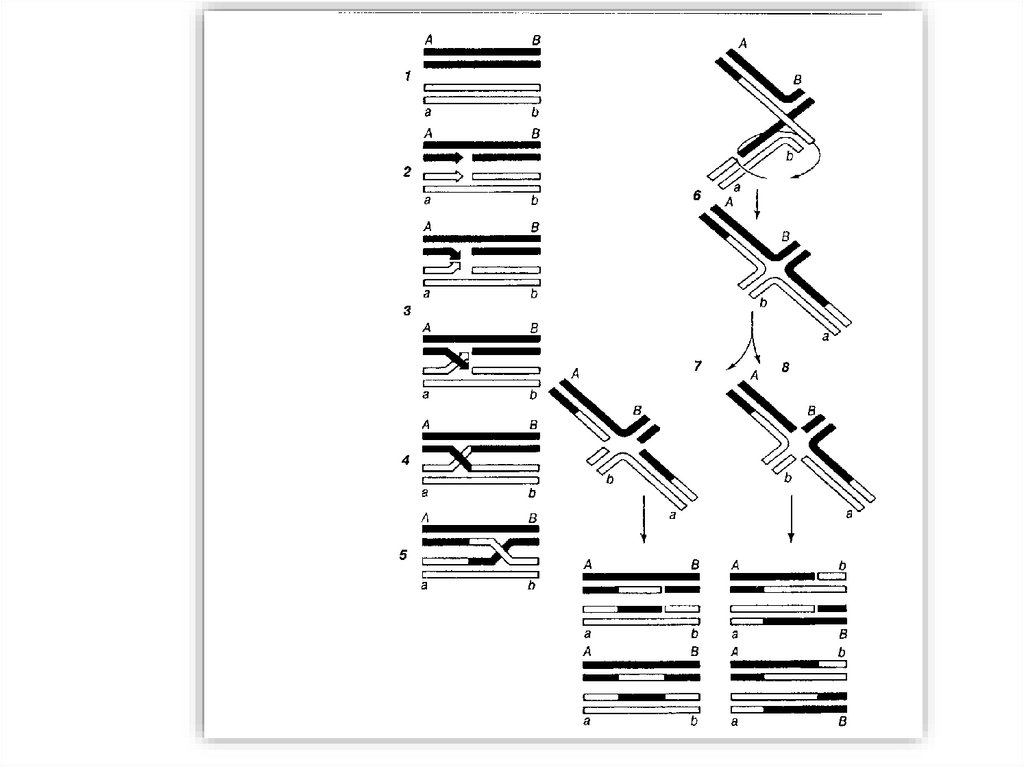
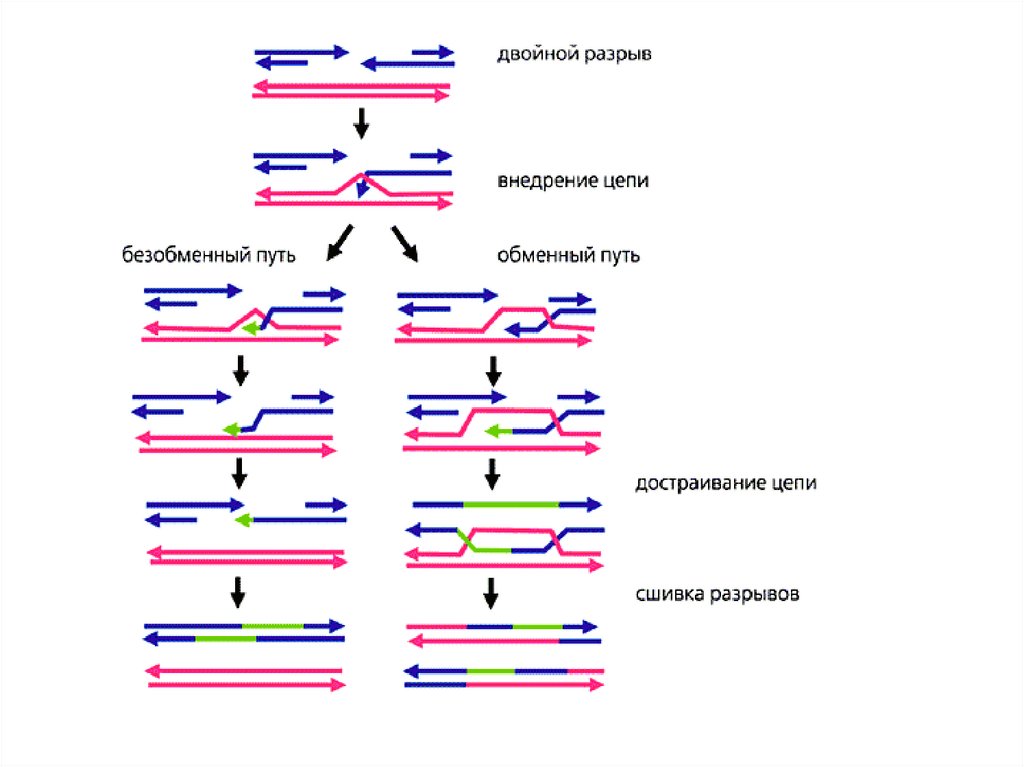
Модели рекомбинацииМодель Холлидея (1964)
Модель Мезельсона-Рэддинга ( у ряда Ascomycetes)
Модель Жостака
4.
5.
6.
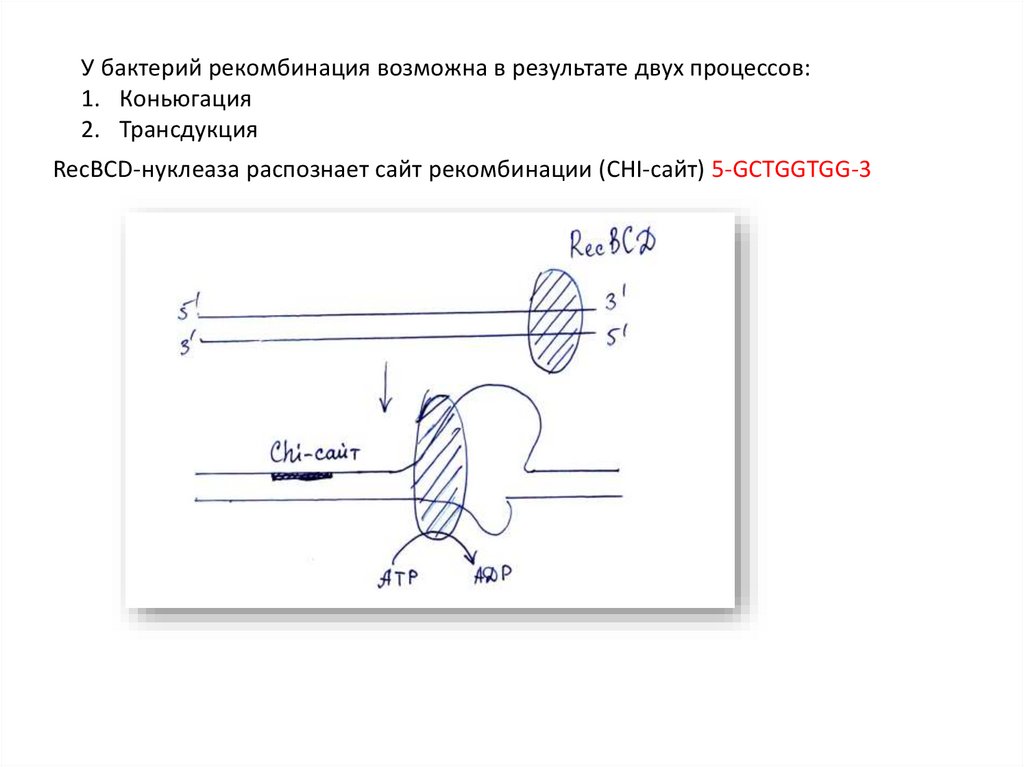
У бактерий рекомбинация возможна в результате двух процессов:1. Коньюгация
2. Трансдукция
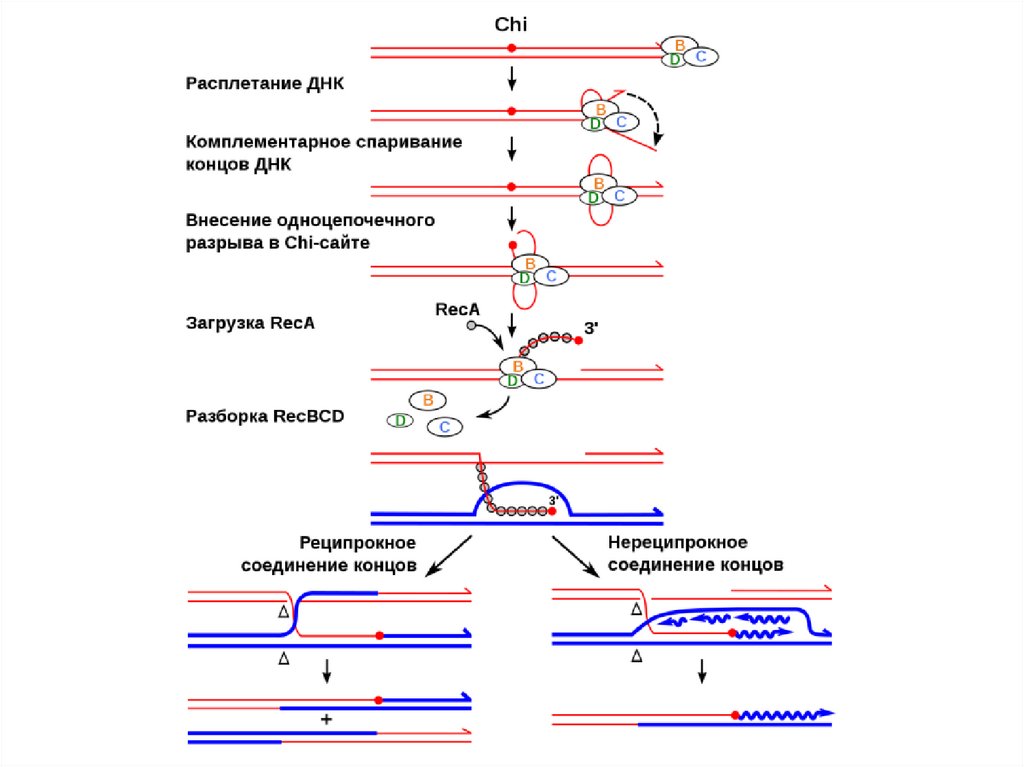
RecBCD-нуклеаза распознает сайт рекомбинации (CHI-сайт) 5-GCTGGTGG-3
7.
RecA-ДНК филамент, два филамента не рекомбинируют друг с другомИзменение конформации двуцепочной ДНК (шаг составляет 18,6 п.о.),
молекула ДНК оказывается растянутой в 1,5 раза
RecA, 38кДа, два домена (первый - связь с переносимой ДНК,
второй домен – взаимодействие с целевой ДНК
Требуется АТФ и одноцепочный фрагмент ДНК
(бреш или свободный конец)
8.
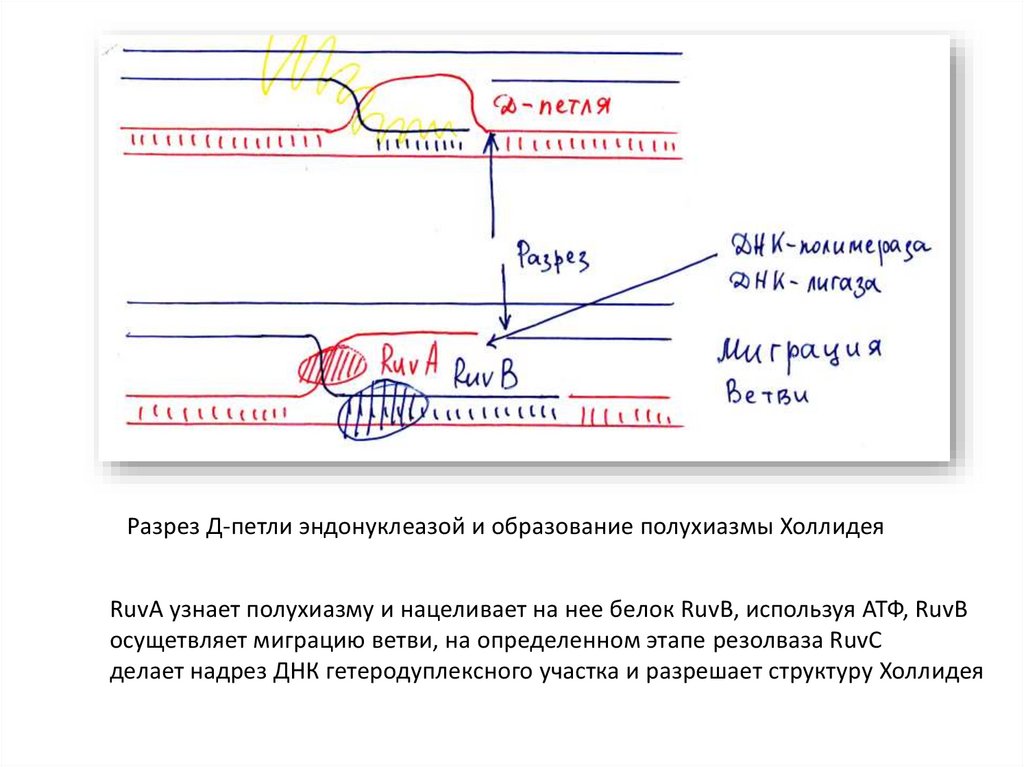
Разрез Д-петли эндонуклеазой и образование полухиазмы ХоллидеяRuvA узнает полухиазму и нацеливает на нее белок RuvB, используя АТФ, RuvB
осущетвляет миграцию ветви, на определенном этапе резолваза RuvC
делает надрез ДНК гетеродуплексного участка и разрешает структуру Холлидея
9.
Происходит в профазу мейоза I(сперматоцит – 24 дня, у женщин начинается на 3-ем месяце
внутриутробного развития и длится до 50-летнего возраста),
постепенная гибель ооцитов:
•У эмбриона 7х106 ооцитов,
•при рождении 2х106 ооцитов
•половое созревание 3х105 ооцитов
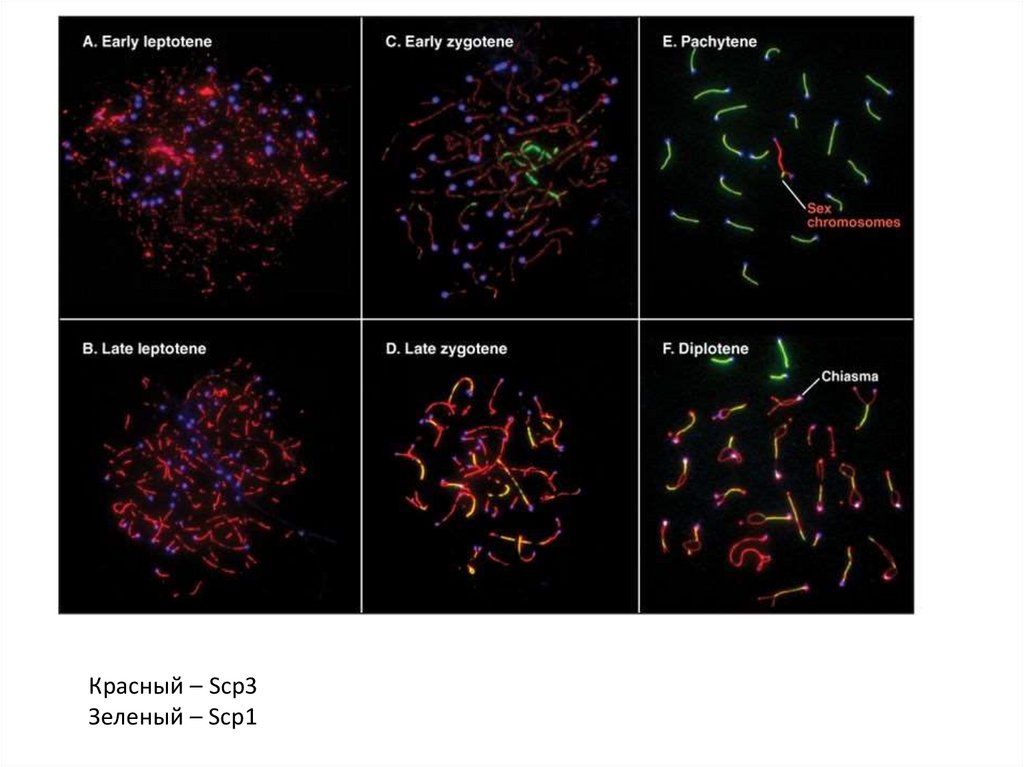
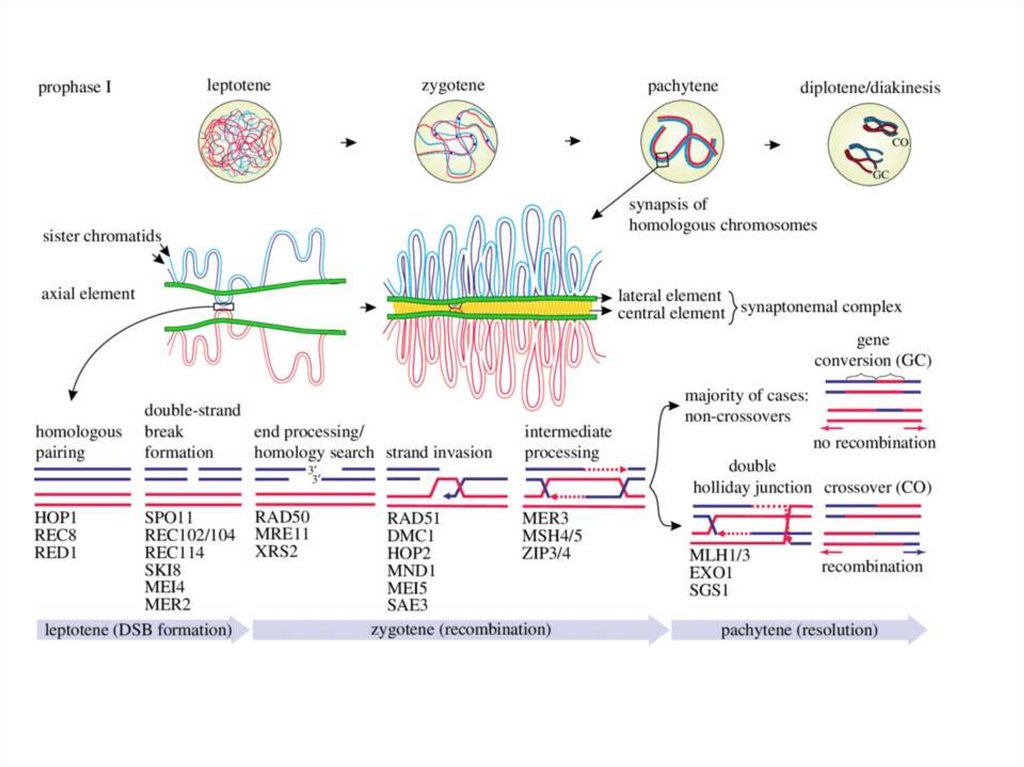
Профаза мейоза:
•Лептотена
•Зиготена
•Пахитена
•Диплотена
•Диакинез
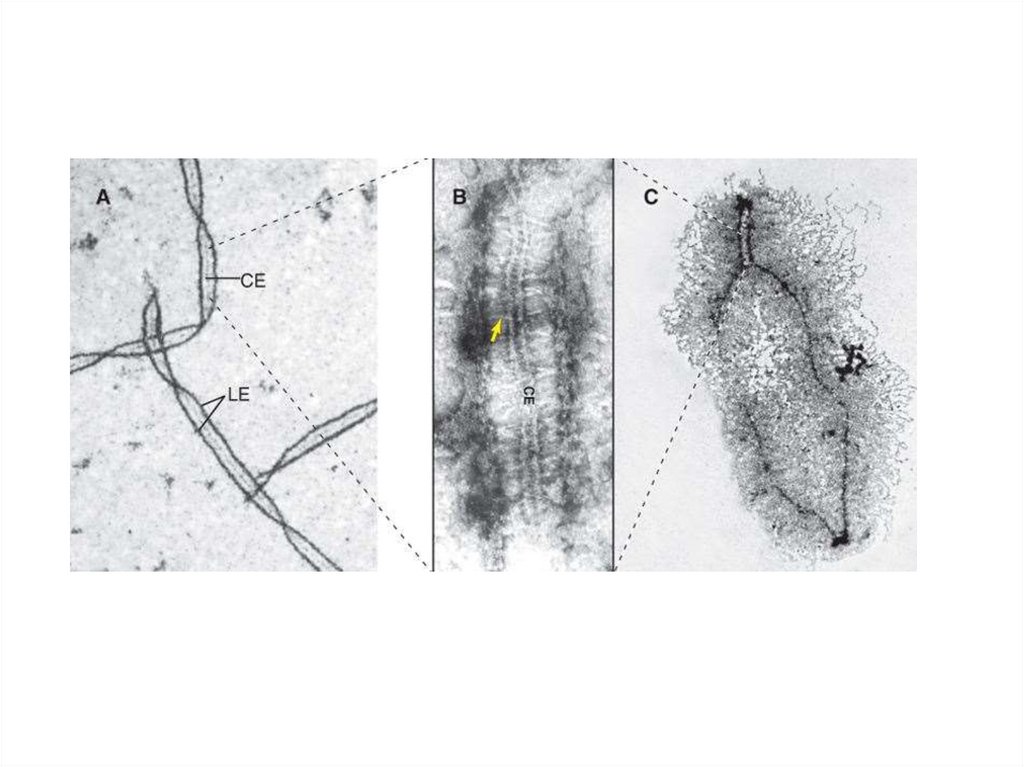
10.
11.
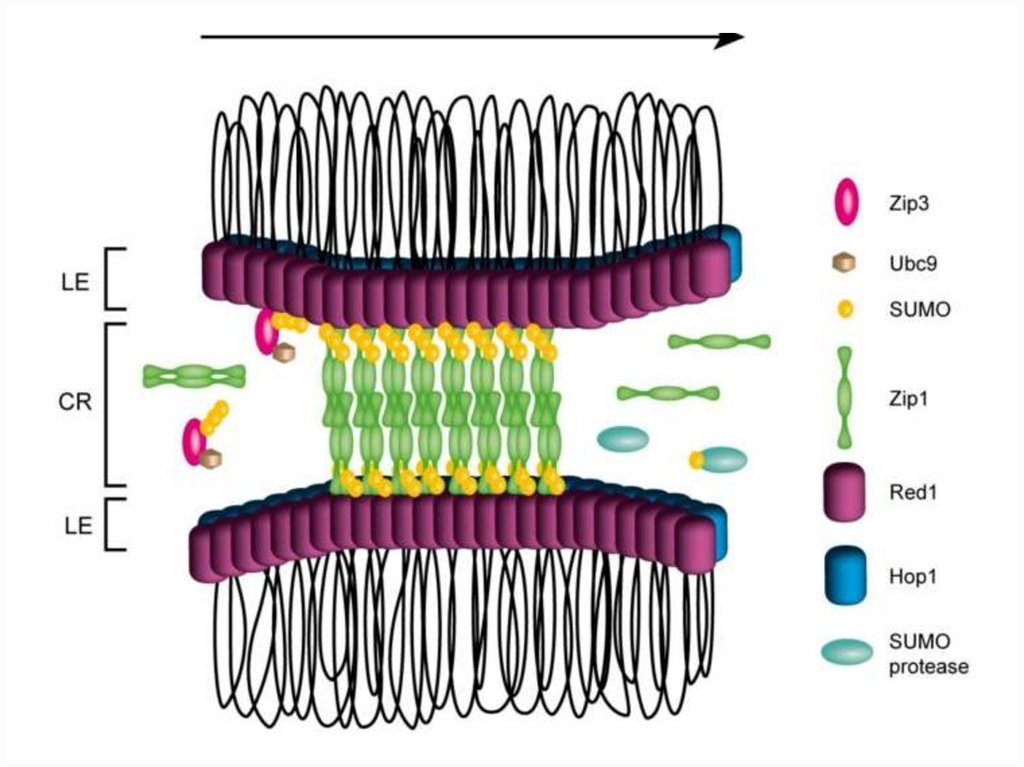
Красный – Scp3Зеленый – Scp1
12.
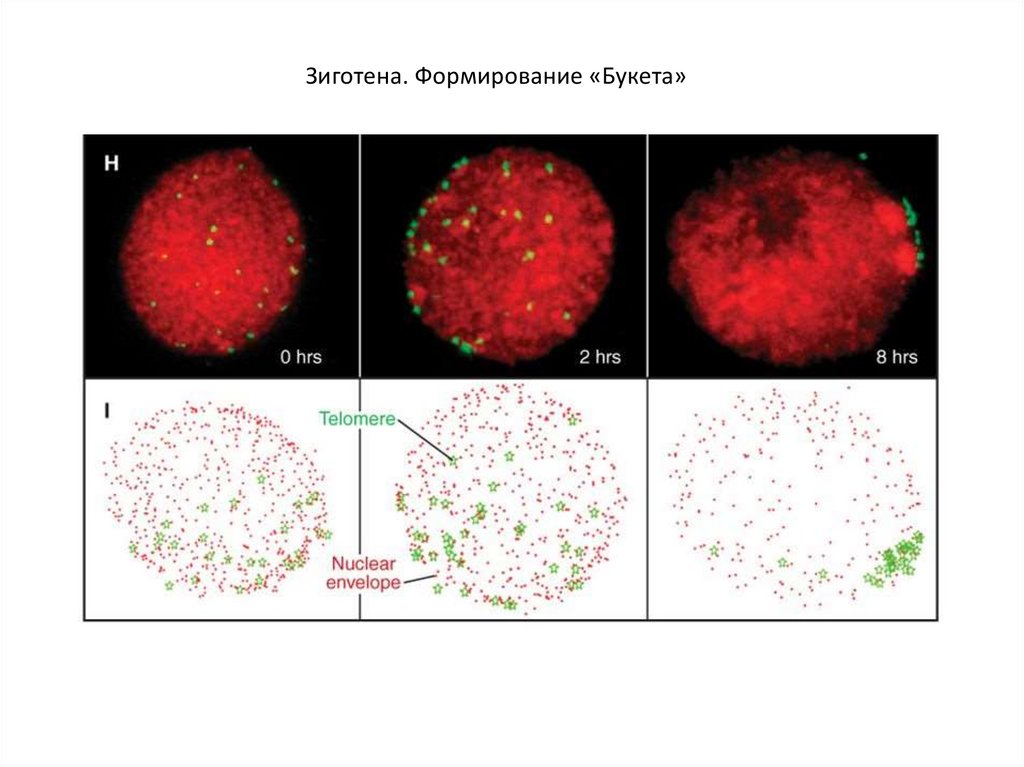
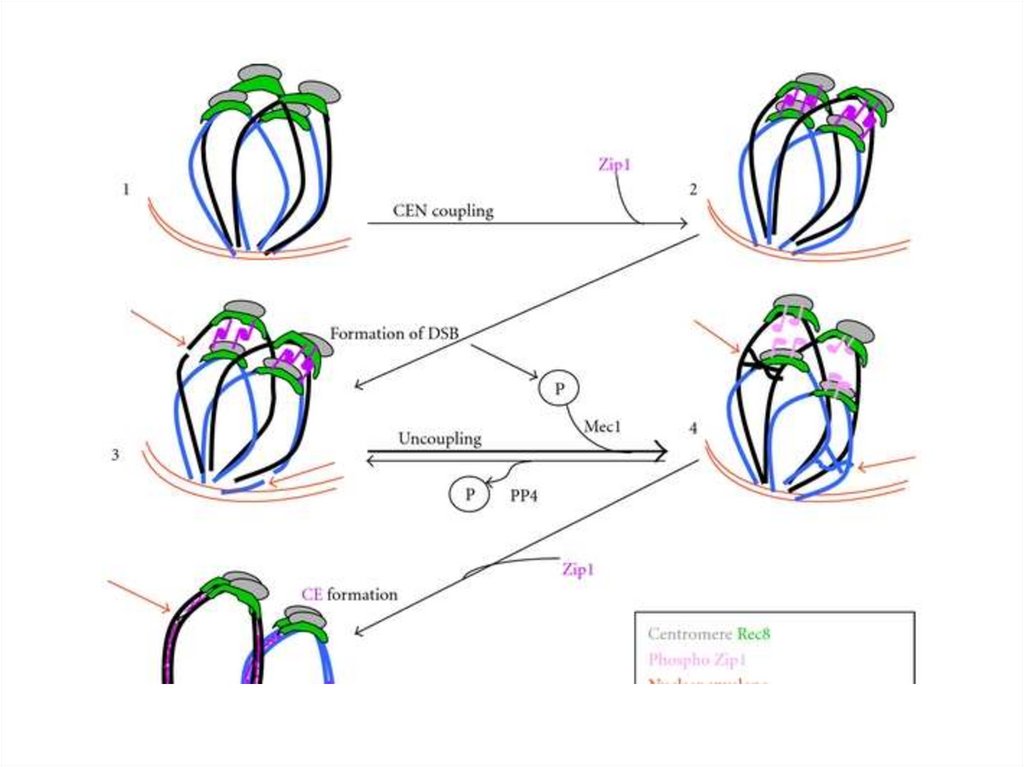
Зиготена. Формирование «Букета»13.
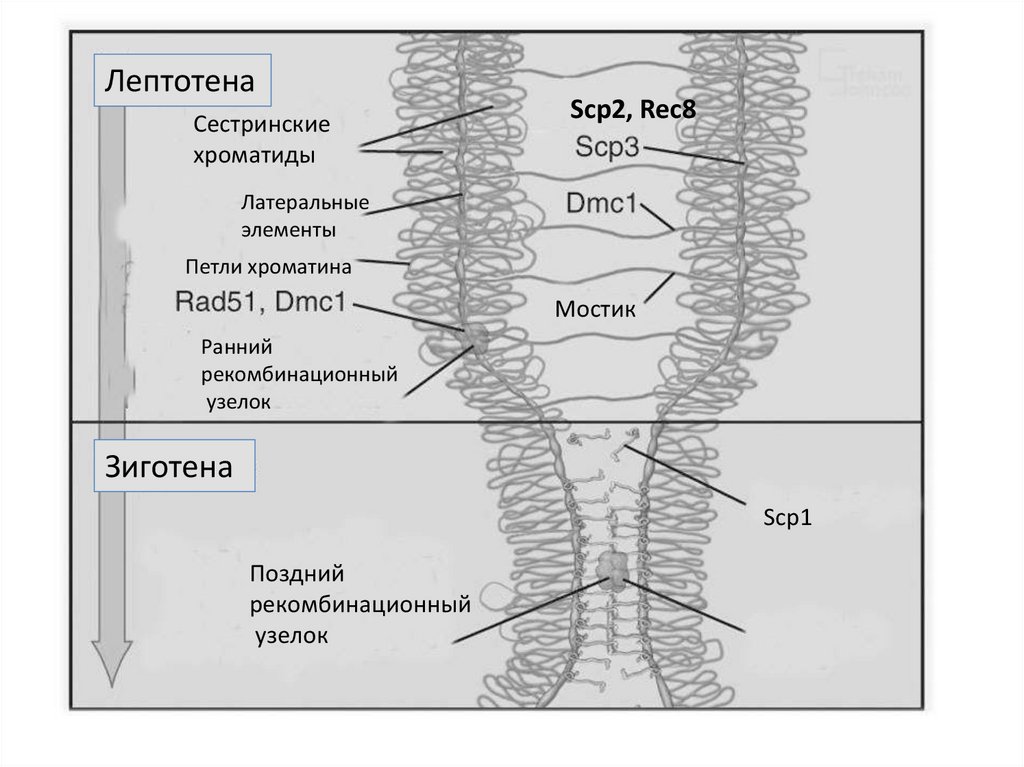
ЛептотенаСестринские
хроматиды
Scp2, Rec8
Латеральные
элементы
Петли хроматина
Мостик
Ранний
рекомбинационный
узелок
Зиготена
Scp1
Поздний
рекомбинационный
узелок
14.
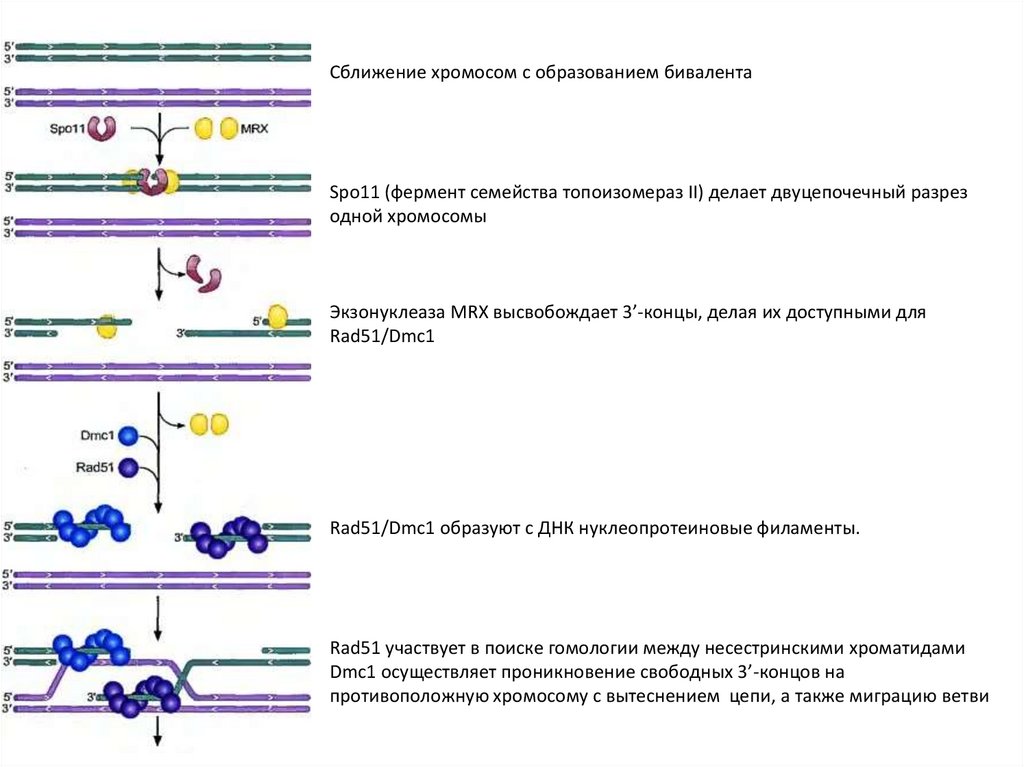
Сближение хромосом с образованием бивалентаSpo11 (фермент семейства топоизомераз II) делает двуцепочечный разрез
одной хромосомы
Экзонуклеаза MRX высвобождает 3’-концы, делая их доступными для
Rad51/Dmc1
Rad51/Dmc1 образуют с ДНК нуклеопротеиновые филаменты.
Rad51 участвует в поиске гомологии между несестринскими хроматидами
Dmc1 осуществляет проникновение свободных 3’-концов на
противоположную хромосому с вытеснением цепи, а также миграцию ветви
15.
16.
17.
18.
19.
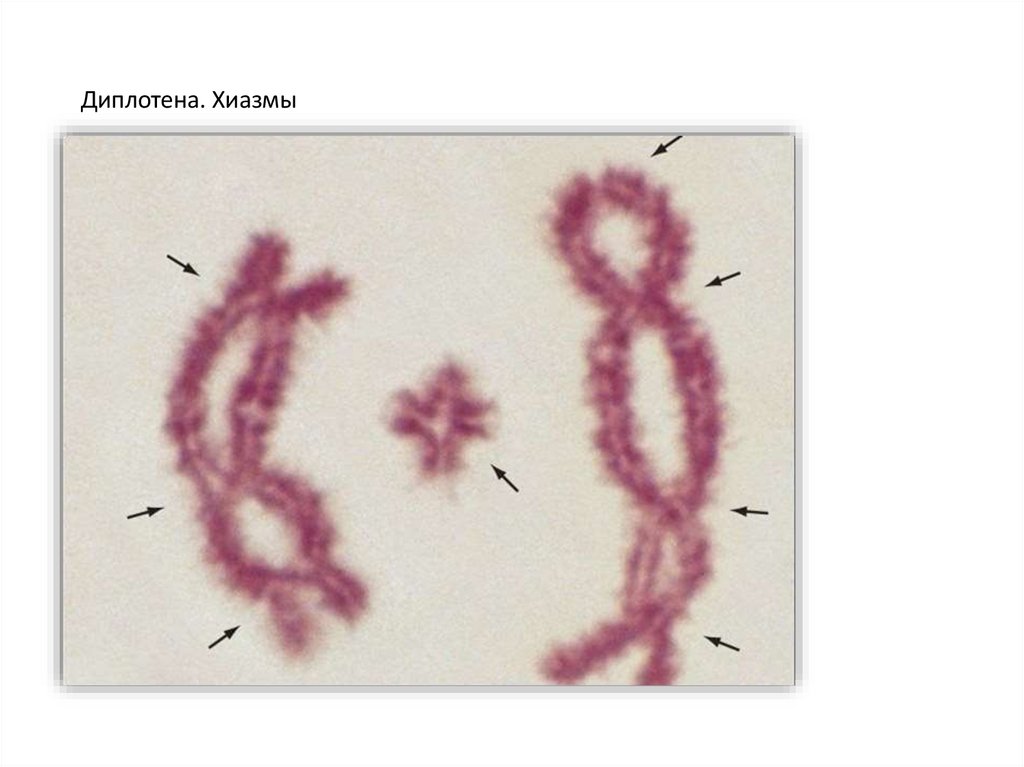
Диплотена. Хиазмы20.
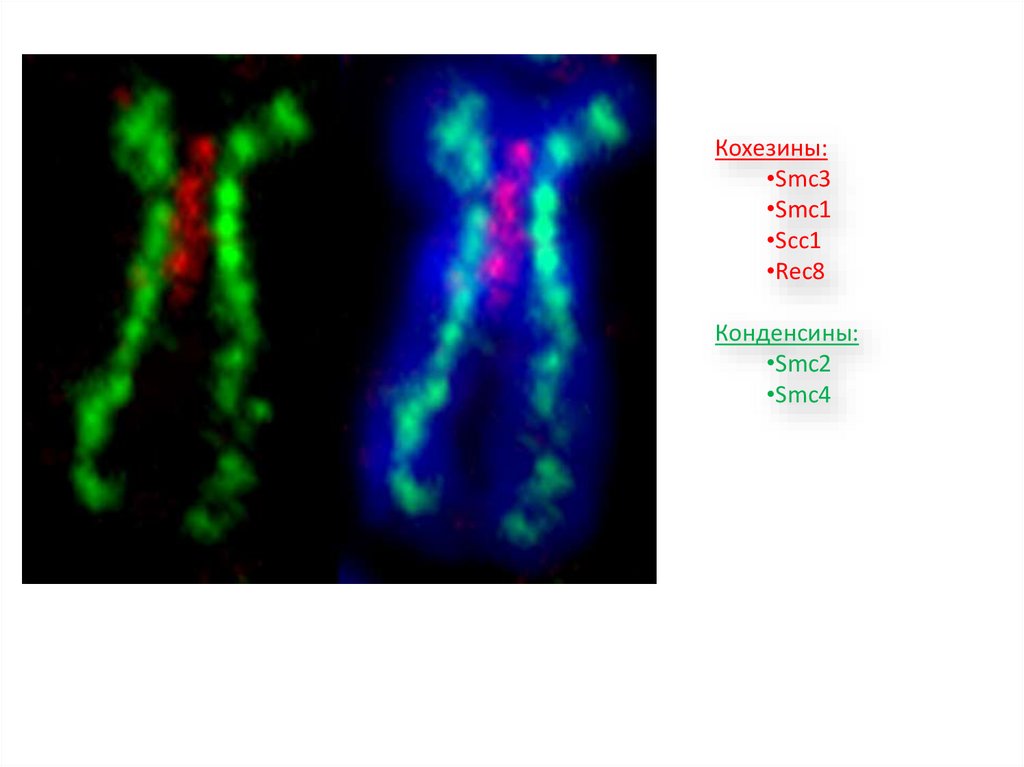
Кохезины:•Smc3
•Smc1
•Scc1
•Rec8
Конденсины:
•Smc2
•Smc4
21.
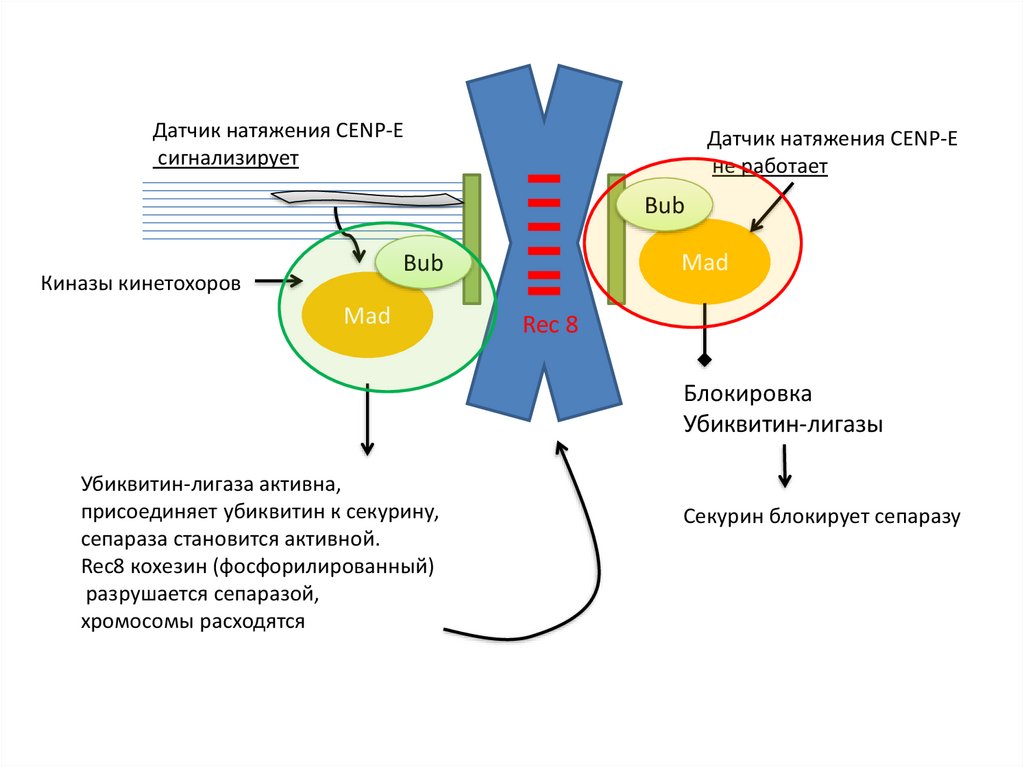
Датчик натяжения CENP-Eсигнализирует
Датчик натяжения CENP-E
не работает
Bub
Mad
Bub
Киназы кинетохоров
Mad
Rec 8
Блокировка
Убиквитин-лигазы
Убиквитин-лигаза активна,
присоединяет убиквитин к секурину,
сепараза становится активной.
Rec8 кохезин (фосфорилированный)
разрушается сепаразой,
хромосомы расходятся
Секурин блокирует сепаразу
22.
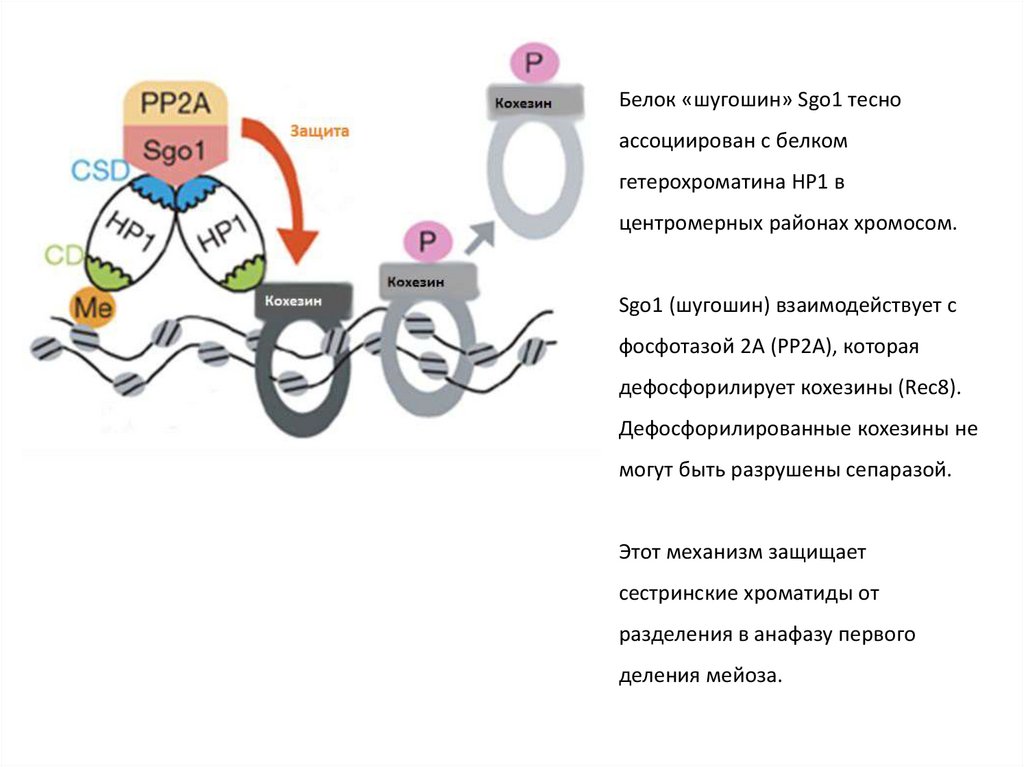
Белок «шугошин» Sgo1 тесноассоциирован с белком
гетерохроматина HP1 в
центромерных районах хромосом.
Sgo1 (шугошин) взаимодействует с
фосфотазой 2А (РР2А), которая
дефосфорилирует кохезины (Rec8).
Дефосфорилированные кохезины не
могут быть разрушены сепаразой.
Этот механизм защищает
сестринские хроматиды от
разделения в анафазу первого
деления мейоза.
23.
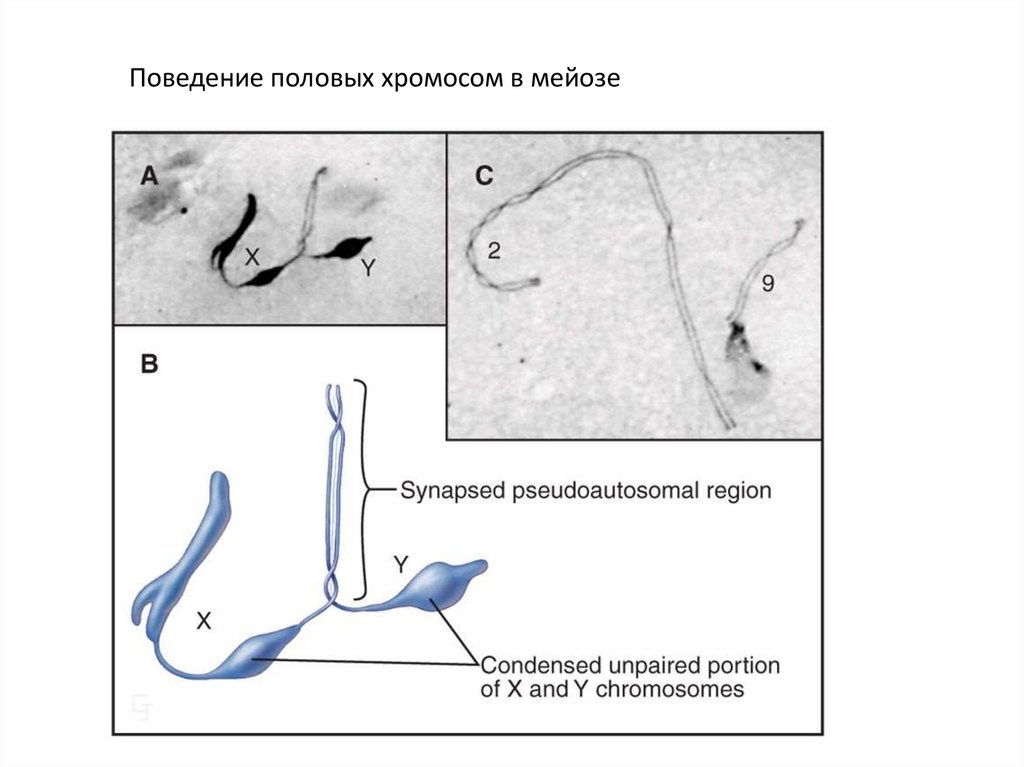
Поведение половых хромосом в мейозе24.
25.
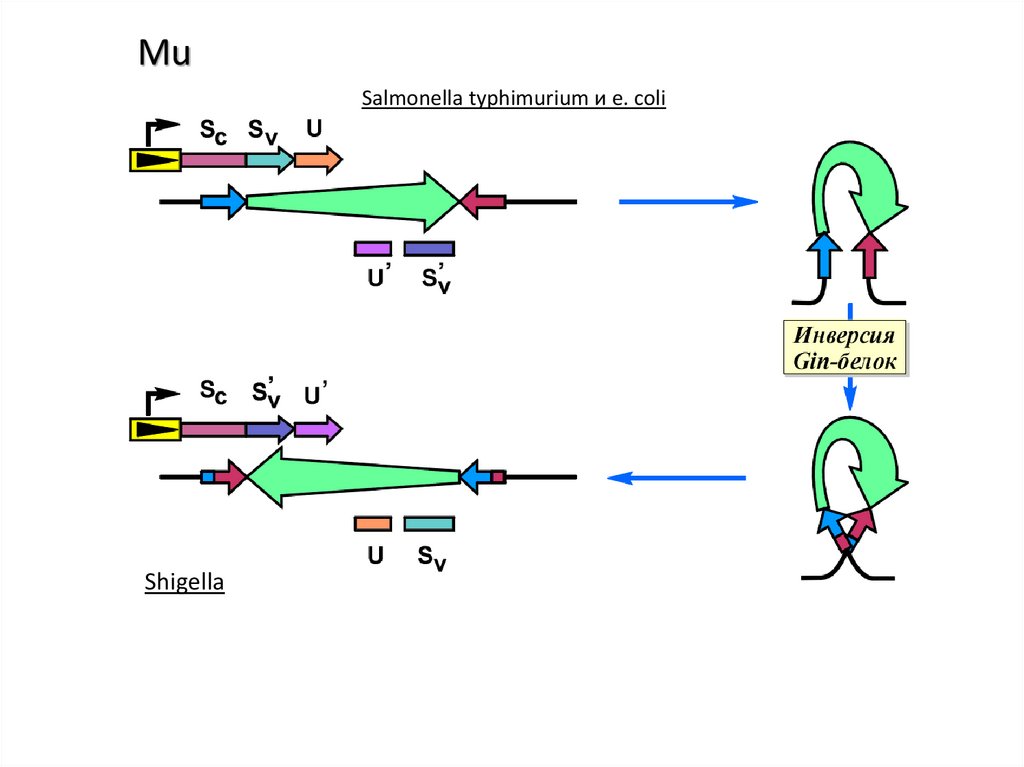
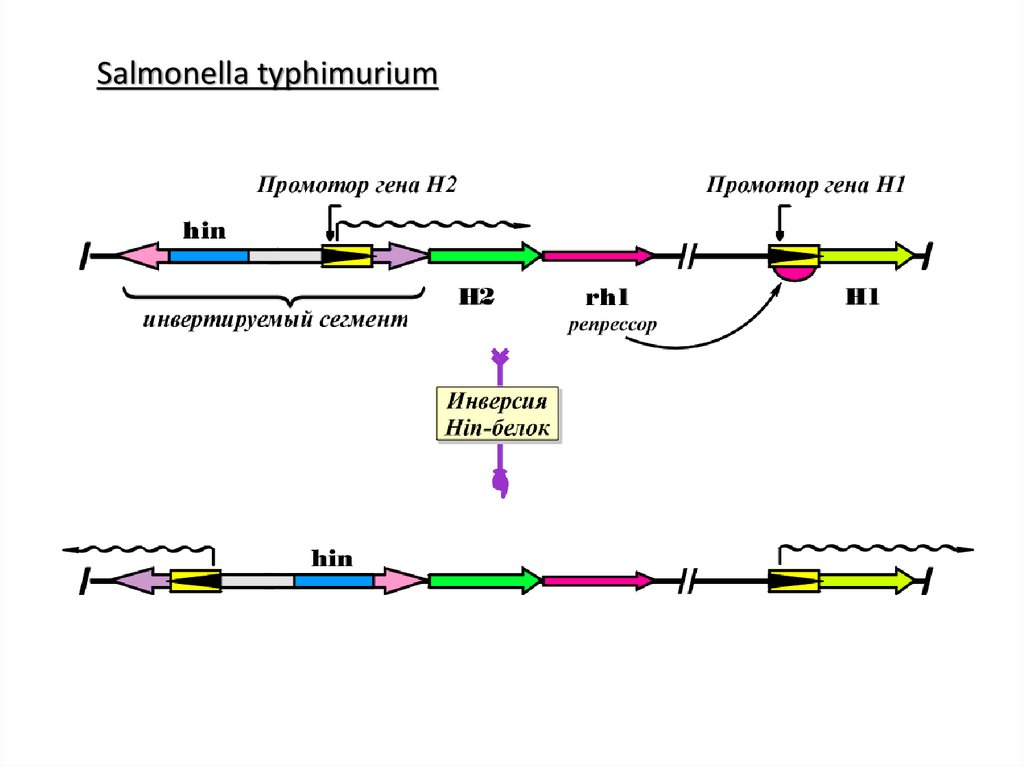
MuSalmonella typhimurium и e. coli
Shigella