




































 biology
biologySimilar presentations:










Обмен углеводов
1.
ОБМЕН УГЛЕВОДОВГликолиз. Пентозофосфатный путь
окисления глюкозы.
2.
ПЛАН ЛЕКЦИИ1. Переваривание и всасывание углеводов в
желудочно-кишечном тракте.
2. Этапы и последовательность реакций аэробного
и анаэробного окисления глюкозы в тканях
(гликолиза).
3. Энергетический эффект аэробного и
анаэробного гликолиза.
4. Пентозофосфатный путь окисления глюкозы в
тканях (окислительный этап, локализация,
значение).
3.
Обмен углеводов в организме человекаскладываются из следующих процессов:
1. Расщепление в желудочно-кишечном тракте
до моносахаридов
поступающих
с
пищей
полисахаридов
и
дисахаридов.
Всасывание
моносахаридов из кишечника в кровь.
2. Синтез и распад гликогена.
3. Анаэробное и аэробное расщепление глюкозы.
В тканях существует два основных пути распада
глюкозы: анаэробный путь гликолиза, который идет
без потребления кислорода и аэробный путь прямого
окисления глюкозы.
4.
4. Пентозофосфатный путь.5. Аэробный метаболизм пирувата, включающий
окислительное декарбоксилирование пирувата и
превращение ацетил-КоА в ЦТК.
6. Глюконеогенез, т. е. образование углеводов из
неуглеводных продуктов, таких как пируват, лактат,
глицерин, аминокислоты.
5.
ОБМЕН УГЛЕВОДОВГлюконеогенез
Распад(гликогенолиз)
и синтез(гликогенез)
гликогена
Гликолиз
Окислительное
декарбоксилирование
пирувата
ЦТК
Пентозофосфатный
путь
6.
Основные функции углеводов:I. Резервная (крахмал, гликоген)
II. Структурная (полисахариды образуют прочный
остов в комплексе с белками и липидами, они входят в
состав биомембран)
III.Защитная (кислые гетерополисахариды выполняют
роль биологического смазочного материала)
IV.Обеспечивают до 70% потребности организма в
энергии.
V. Являются источниками углеродов, который
необходим для синтеза ряда соединений ( белков,
липидов и т.д.).
7.
Переваривание углеводов ворганизме
Источником углеводов для организма служат
углеводы пищи - крахмал, сахароза и лактоза. Кроме
того, глюкоза может образовываться в организме из
аминокислот, глицерина.
Углеводы пищи в пищеварительном тракте
распадаются на мономеры. В переваривании
принимают участие гидролазы. Специфические
гидролазы: мальтаза, сахараза, лактаза
вырабатываются клетками кишечника и содержатся в
кишечном соке.
8.
Продукты полного переваривания углеводов –глюкоза, галактоза и фруктоза – через клетки
кишечника поступают в кровь. При всасывании из
кишечника в кровь моносахариды проникают через
клеточные мембраны путем облегченной диффузии
и с помощью активного транспорта. Активный
транспорт обеспечивает перенос моносахаридов
против градиента концентрации, и поэтому может
функционировать тогда, когда концентрация
глюкозы или галактозы в кишечнике невелика.
Важнейшие сахара через воротную вену
проникают в печень, где идет превращение
фруктозы, галактозы и глюкозы.
9.
ВСАСЫВАНИЕ ПРОДУКТОВ ПЕРЕВАРИВАНИЯУГЛЕВОДОВ
Гексозы (глю, гал,
фру) в энтероциты всасываются
против градиента концентрации путем:
1. облегченной диффузии (белки-переносчики)
2. активного Na+ -зависимого транспорта (глю и гал)
(затрата энергии, которую обеспечивает работа Na+, K+-АТФ-азы)
Пентозы (рибоза, дезоксирибоза) всасываются по градиенту
концентрации
Из энтероцитов в кровь (воротная вена) гексозы попадают
путем облегченной диффузии
Из крови глюкоза поступает в клетки тканей путем облегченной
диффузии при участии специальных ферментов ГЛЮТ 1-2-3-4-5
(Глют-4 – инсулинозависимый в миоцитах и адипоцитах)
10.
ГликолизГликолиз (от греч. glykys – сладкий, lysys –
распад ) – один центральных путей катаболизма
глюкозы.
В процессе гликолиза происходит расщепление
шестиуглеродной молекулы глюкозы на две
трехуглеродные молекулы пирувата .
Основное значение:
1. производство
универсального продукта
окисления
2. обеспечение энергией клетки в условиях
гипоксии.
11.
Стадии гликолиза:Подготовительная стадия, которая состоит из
пяти этапов. Продуктом первой стадии гликолиза
является глицеральдегид-3-фосфат. Подготовительная
стадия гликолиза служит для того, чтобы превратить
углеродные цепочки всех метаболизируемых гексоз в
один общий продукт – глицеральдегид-3-фосфат.
Вторая стадия гликолиза также состоит из пяти
ферментативных реакций; на этой стадии энергия,
высвобождающаяся при превращении двух молекул
глицеральдегид-3-фосфата в две молекулы пирувата,
запасается в виде четырех молекул АТФ.
12.
Гликолиз включает превращениятрех разных типов:
1. Распад углеродного скелета глюкозы с
образованием пирувата.
2. Фосфорилирование
АДФ
высокоэнергетическими фосфорилированными
соединениями с образованием АТФ .
3. Перенос водородных атомов или электронов.
Ферменты, катализирующие гликолиз,
локализованы в цитозоле.
13.
Реакции гликолизаI стадия (подготовительная)
1. Фосфорилирование глюкозы: реакция протекает необратимо (I),
катализируется
регуляторным
ферментом
гексокиназой
(глюкокиназой в печени и поджелудочной железе) и требует
затраты АТФ.
2. Образование
глюкозо-6-фосфата
в
гексокиназной
реакции
сопровождается
освобождением
значительного количества свободной энергии системы и считается
необратимым процессом.
CH2OH
гексокиназа
глюкокиназа
O
2+
АТФ
глю
Mg
CH2OPO3H2
O
АДФ
глю-6-ф
14.
2. Превращение глю-6-ф в фру-6-ф.Вторая реакция необходима для выведения еще одного атома
углерода из кольца для его последующего фосфорилирования
(фермент фосфоглюкоизомераза). В результате образуется
фруктозо-6-фосфат.
CH2OPO3H2
O
CH2OPO3H2
фосфоглюкоизомераза
O
OH
2+
Mg
Глю-6-ф
CH2OH
Фру-6-ф
15.
3. Фосфорилирование фру-6-ф во фру-1,6-фф.CH 2 OPO 3 H 2
O
фосфофруктокиназа
OH
CH2OPO3H2
O
2+
OH
Mg
АТФ
CH 2 OH
Фру-6-ф
АДФ
CH2OPO3H2
фру-1,6-диф
Фосфофруктокиназа является регуляторным и аллостерическим
ферментом. Эта стадия требует затраты АТФ. Это необратимая реакция (II)
гликолиза.
В третьей реакции происходит фосфорилирование (присоединение
остатка ортофосфорной кислоты) фруктозо-6-фосфата с образованием
фруктозо-1,6-дифосфата. При этом затрачивается еще одна молекула АТФ
(уже вторая) - это вторая пусковая реакция гликолиза. Она идет в
присутствии Mg2+ и является необратимой, так как сопровождается
масштабным уменьшением свободной энергии.
16.
4. Расщепление фру-1,6-фф на фосфотриозы.O
CH2OPO3H2
O
OH
OH
C
альдолаза
H
CH
CH2OPO3H2
фру-1,6-диф
CH2
OH
+
CH2OPO3H2
глицеральдегид-3-ф
5%
C
O
CH2OPO3H2
дигидроксиацетонфосфат
95%
В
дальнейших
превращениях
принимает
участие
глицеральдегид-3-ф, который образуется в результате изомеризации
дигидроксиацетонфосфата:
5. дигидроксиацетонфосфат
глицеральдегид-3-фосфат
На первой стадии из одной молекулы глю образуется две молекулы
глицеральдегид-3-фосфата, которые участвует в дальнейших превращениях
17.
На второй стадии гликолиза запасается энергия.1.
Окисление
глицеральдегид-3-фосфата
до
кислоты
и
присоединение к ней фосфорной кислоты приводит к образованию
макроэргического соединения 1,3-дифосфоглицериновой кислоты и
НАДН.
O
O
глицеральдегидфосфатдегидрогеназа
C
H
2
CH
C
О OPO
~ PO3H
H2
3
2
OH
CH2OPO3H2
глицеральдегид-3-ф
+
2НАД Фн
+
2НАДН+Н
CH
2
OH
CH2OPO3H2
1,3-дифосфоглицерат
18.
2. Образование 3-фосфоглицерата.Субстратное фосфорилирование
2АДФ
O
2АТФ
C
Mg
О~PO
OPO3H
22
3H
2
CH
COOH
2+
OH
CH2OPO3H2
фосфоглицераткиназа
2
CH
OH
CH2OPO3H2
3-фосфоглицерат
В седьмой реакции фосфорильная группа 1,3-бифосфоглицерата
переносится на AДФ, в результате чего образуется АТФ
(напоминаем, что следует иметь в виду две параллельные цепи
реакций, с участием двух молекул триоз, образовавшихся из одной
молекулы гексозы, следовательно, синтезируется не одна, а две
молекулы АТФ)
19.
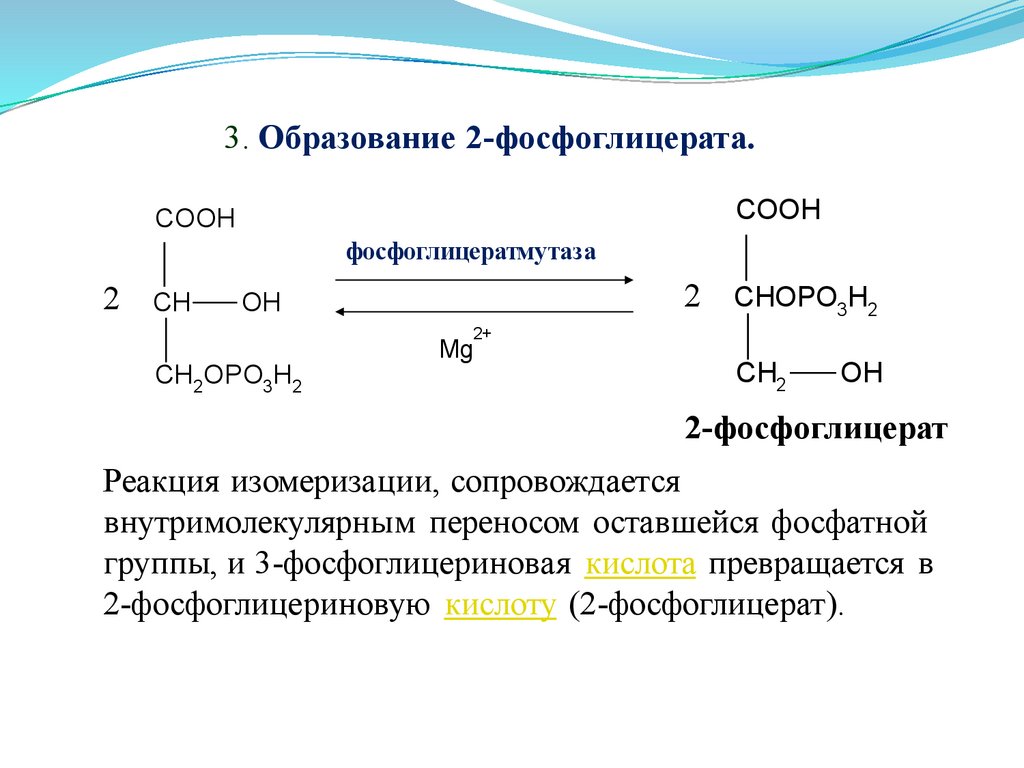
3. Образование 2-фосфоглицерата.COOH
COOH
фосфоглицератмутаза
2
CH
2 CHOPO3H2
OH
2+
Mg
CH2OPO3H2
CH2
OH
2-фосфоглицерат
Реакция изомеризации, сопровождается
внутримолекулярным переносом оставшейся фосфатной
группы, и 3-фосфоглицериновая кислота превращается в
2-фосфоглицериновую кислоту (2-фосфоглицерат).
20.
4. Образование фосфоенолпирувата.COOH
COOH
2
CHOPO3H2
енолаза -Н2О
+Н2О
CH2
OH
2 C
О~
PO3HH2
OPO
3
2
CH2
фосфоенолпируват
(высокоэнергетическое соединение)
Девятая реакция – Реакция енолизации,
сопровождается внутримолекулярными окислительновосстановительными процессами, в результате которых
образуется фосфоенолпируват
(фосфоенолпировиноградная кислота) с
высокоэнергетической связью во втором атоме углерода
и отщепляется молекула воды.
21.
5. Образование пирувата -необратимая реакция гликолиза (III)Субстратное фосфорилирование
Заключается в переносе пируваткиназой макроэргического фосфата с
фосфоенолпирувата на АДФ с образованием пировиноградной кислоты
и АТФ, высокоэнергетичная.
COOH
2
C
CH2
OPO
О~PO3H
3H22
2АДФ
2+
Mg
2АТФ
COOH
2 C
O
пируваткиназа
C H3
пируват
22.
Дальше процесс идет в зависимости от наличия илиотсутствия кислорода в клетке:
При анаэробных условиях электроны, пришедшие сначала
+
от глицеральдегид-3-фосфата к НАД , переносятся в форме
НАД·Н + Н+ на пируват, который восстанавливается до
лактата.
CO OH
COOH
2
C
CH3
пируват
O
лактатдегидрогеназа
2НАД·Н + Н+
2НАД
2
CH
CH 3
OH
лактат
23.
Последняя реакция бескислородногоокисления глюкозы, одиннадцатая –
образование молочной кислоты из пирувата
под действием лактатдегидрогеназы.
Важно то, что эта реакция осуществляется
только в анаэробных условиях.
Эта реакция необходима клетке, так как
НАДН, образующийся в 6-й реакции, в
отсутствие кислорода не может окисляться
в митохондриях.
24.
Суммарная реакцияанаэробного гликолиза
При анаэробном гликолизе образуется 4 молекулы
АТФ, но клетке остается только 2 молекулы, т. к. 2
молекулы
АТФ
затрачивается
в
процессе
фосфорилирования
в
первой
подготовительной
(энегопотребляющей) стадии гликолиза.
Глю
+
2АДФ
+
2Фн
2лактат + 2АТФ
25.
Пути превращения пируватаглю
COOH
2
CH
COOH
NH2
+ NH3
O
C
2
COOH
аланин
COOH
ЛДГ
НАД·Н + Н+
2
2 CH3
OH
CH3
лактат
COOH
пируват
Окислительное
декарбоксилирование
CH
дых. цепь - 6 АТФ
+2НАД·Н + Н+
ПДГ
O
C
SKoA
+
ЦТК – 3 НАДН+Н - 9АТФ
ФАДН2 - 2АТФ
ГТФ
- 1АТФ
12АТФ
26.
Конечным продуктом аэробного гликолиза являетсяпируват, а энергетический баланс складывается из 2
молекул АТФ образовавшихся в результате субстратного
фосфорилирования и остается еще 2 молекулы
восстановленного НАД·Н + Н +, от концентрации которого
зависит скорость процесса. Для продолжения процесса
необходим сброс Н + на ферменты дыхательной цепи, но
сама молекула НАД·Н + Н + через мембрану митохондрий
проникнуть не может, для этого используются переносчики
и перенос осуществляется с помощью 2-х механизмов:
1. Глицерофосфатный челночный механизм;
2. Малат – аспартатный челночный механизм;
27.
Глицерофосфатный челночный механизмЦитоплазма
OH
CH2
2
C
Митохондрии
O
OH
CH2
2
C
КоQ
O
Цв
CH2OPO3H2
CH2OPO3H2
дигидроксиацетонфосфат
АТФ
ФАДН2
2НАД·Н + Н+
2НАД
ФАД+
+
CH2
CH
OH
CH
OH
Ца/а3
АТФ
OH
2
2
CH2
Цс
O2
OH
CH2OPO3H2
альфа-глицеролфосфат
CH2OPO3H2
2ФАДН2
4АТФ
28.
Глицеролфосфатный челночный механизмГлавными ферментами глицеролфосфатного челнока являются
изоферменты глицерол-3-фосфат-дегидрогеназы –
цитоплазматический и митохондриальный. Они отличаются своими
коферментами: у цитоплазматической формы – НАД, у
митохондриальной – ФАД.
В цитозоле метаболиты гликолиза – дигидооксиацетонфосфат и НАДН
образуют глицерол-3-фосфат, поступающий в митохондрии. Там он
окисляется с образованием ФАДН2. Далее ФАДН2 направляется в
дыхательную цепь и используется для получения энергии. Таким
образом, в результате действий челнока цитозольный НАДН+H+ как бы
"превращается" в митохондриальный ФАДН2.
В действительности цитозольный глицерол-3-фосфат не проникает в
матрикс, так как митохондриальная глицерол-3-фосфат-дегидрогеназа
расположена на внешней стороне внутренней митохондриальной
мембраны. Она обеспечивает перенос атомов водорода от глицерол-3фосфата на ФАДН2 и дальнейшую передачу их на коэнзим Q
дыхательной цепи.
29.
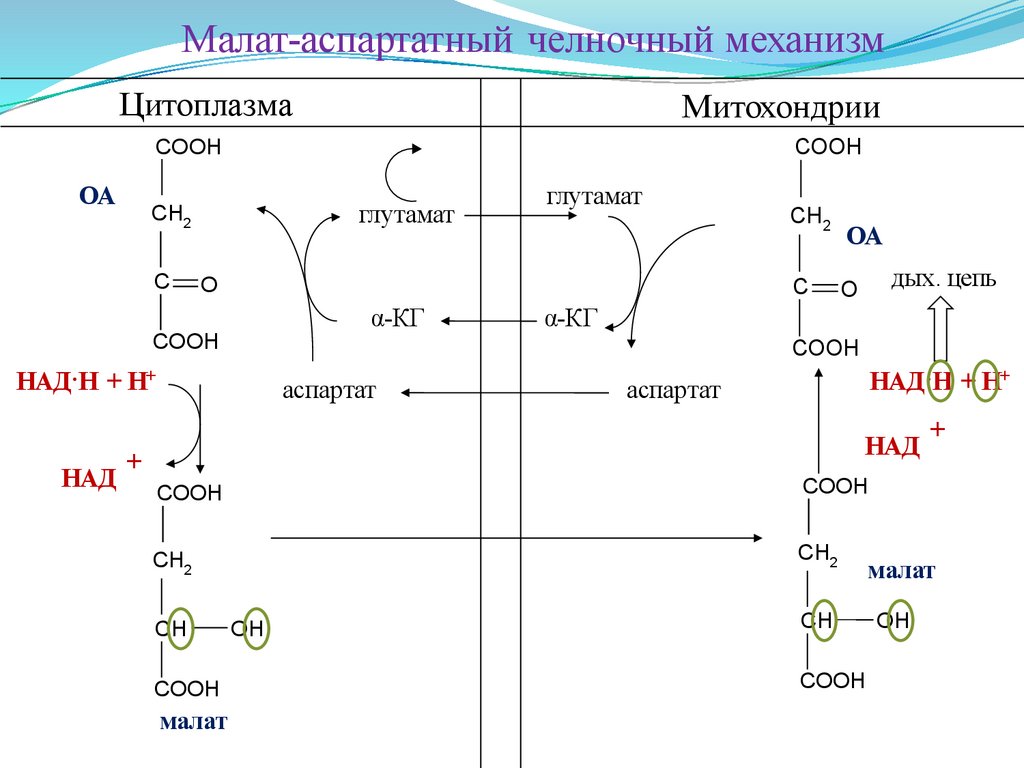
Малат-аспартатный челночный механизмЦитоплазма
Митохондрии
COOH
ОА
COOH
глутамат
CH2
C
глутамат
O
C
α-КГ
ОА
дых. цепь
O
α-КГ
COOH
COOH
НАД·Н + Н+
НАД
CH2
аспартат
НАД·Н + Н+
аспартат
НАД
+
COOH
COOH
CH2
CH2
CH2
COOH
малат
OH
CH2
COOH
+
малат
OH
30.
Малат-аспартатный челночный механизмГлавными
ферментами
этого
челнока
являются
изоферменты
малатдегидрогеназы
–
цитоплазматический
и
митохондриальный. Он является распространенным по всем тканям.
Этот механизм более сложен: постоянно идущие в цитоплазме при участии
фермента аспартатаминотрансферазы (АСТ) реакции трансаминирования
аспарагиновой кислоты с α-кетоглутаратом поставляют оксалоацетат,
который под действием цитозольного пула малатдегидрогеназы и за счет
"гликолитического"
НАДН
восстанавливается
до
яблочной
кислоты (малата).
Последняя антипортом с α-кетоглутаратом проникает в митохондрии и,
являясь метаболитом ЦТК, окисляется в оксалоацетат с образованием
НАДН. Так как мембрана митохондрий непроницаема для оксалоацетата, то
он при помощи аспартатаминотрансферазы трансаминируется до
аспарагиновой кислоты, которая в обмен на глутамат выходит в цитозоль.
Таким образом, атомы водорода от цитозольного НАДН перемещаются в
состав митохондриального НАДН.
31.
Пентозофосфатный путь окисления глюкозы (печень, жироваяткань, эритроциты, молочная железа, надпочечники)
А. ОКИСЛИТЕЛЬНЫЙ ЭТАП
В окислительной части пентозофосфатного пути глюкозо-6-фосфат
подвергается окислительному декарбоксилированию, в результате
которого образуются пентозы. Этот этап включает 2 реакции
дегидрирования.
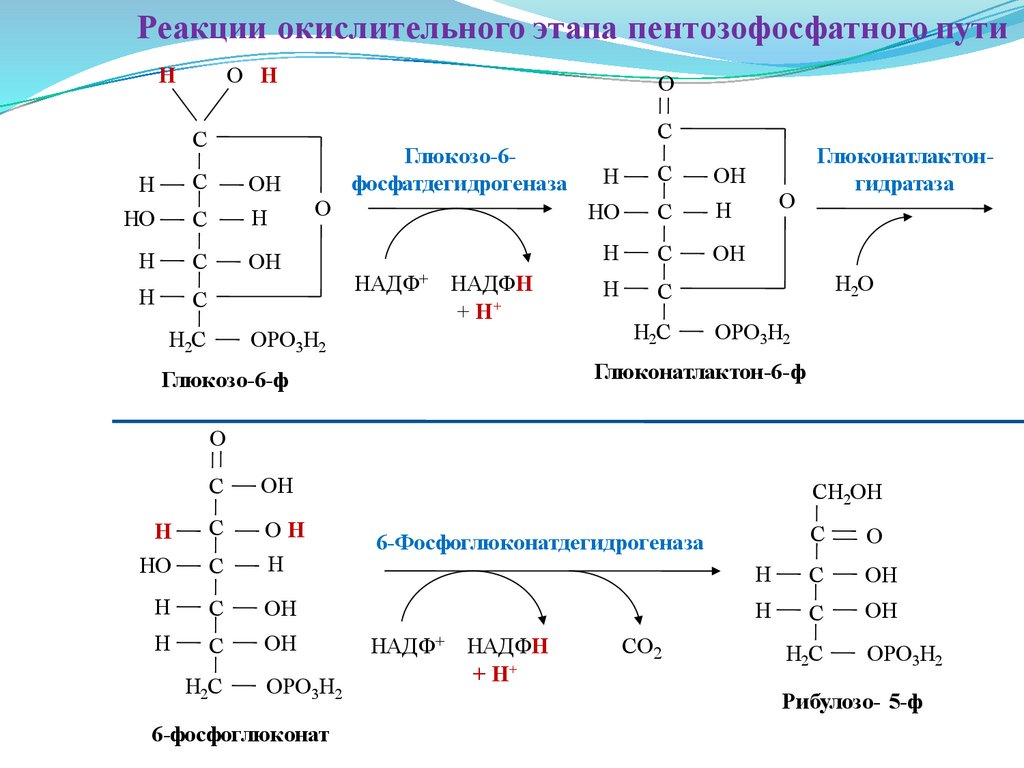
Первая реакция дегидрирования — превращение глюкозо-6-фосфата в
глюконолактон-6- фосфат — катализируется NADP+-зaвиcимoй глюкозо6-фосфатдегидрогеназой и сопровождается окислением альдегидной
группы у первого атома углерода и образованием одной молекулы
восстановленного кофермента NАDРH.
Далее глюконолактон-6-фосфат быстро превращается в 6-фосфоглюконат
при участии фермента глюконолактонгидратазы.
Фермент 6-фосфоглюконатдегидрогеназа катализирует вторую реакцию
дегидрирования окислительной части, в ходе которой происходит также
и декарбоксилирование. При этом углеродная цепь укорачивается на
один атом углерода, образуется рибулозо-5-фосфат и вторая молекула
гидрированного NАDРН.
32.
Восстановленный НАДФН ингибирует первый ферментокислительного этапа пентозофосфатного пути — глюкозо-6фосфатдегидрогеназу. Превращение в НАДФН окисленное
состояние НАДФ + приводит к ослаблению ингибирования
фермента. При этом скорость соответствующей реакции
возрастает, и образуется большее количество НАДФН.
Суммарное уравнение окислительного
этапа пентозофосфатного пути можно представить в виде:
Глю-6-ф+
2НАДФ+
+Н2О
Рибозо-5-ф + 2НАДФН+ Н+2СО2 +2Н+
33.
Реакции окислительного этапа служат основнымисточником НАДФН в клетках. Гидрированные
коферменты снабжают водородом биосинтетические
процессы, окислительно-восстановительные реакции,
включающие защиту клеток от активных форм кислорода.
НАДФН как донор водорода участвует в анаболических
процессах, например, в синтезе холестерина. Это
источник восстановительных эквивалентов для цитохрома
Р450, катализирующего образование гидроксильных групп
при синтезе стероидных гормонов, жёлчных кислот, при
катаболизме лекарственных веществ и других
чужеродных соединений.
34.
Высокая активность фермента глюкозо-6фосфатдегидрогеназы обнаружена вфагоцитирующих лейкоцитах, где НАДФН -оксидаза
использует восстановленный НАДФН для
образования супероксидного иона из молекулярного
кислорода. Супероксидный ион генерирует другие
активные формы кислорода, под действием которых и
повреждаются молекулы ДНК, белков, липидов
бактериальных клеток. Синтез жирных кислот из
углеводов в печени является основным путём
утилизации НАДФН и обеспечивает регенерацию
окисленной формы НАДФ.
35.
Реакции окислительного этапа пентозофосфатного путиН
О Н
О
С
Н
С
ОН
НО
С
Н
Н
С
ОН
Н
С
Н2С
Глюкозо-6фосфатдегидрогеназа
О
НАДФ+
НАДФН
+ Н+
ОРО3Н2
С
Н
С
ОН
НО
С
Н
Н
С
ОН
Н
С
Н2С
О
Глюконатлактонгидратаза
Н2О
ОРО3Н2
Глюконатлактон-6-ф
Глюкозо-6-ф
О
С
ОН
Н
С
ОН
НО
С
Н
Н
С
ОН
Н
С
ОН
Н2С
ОРО3Н2
6-фосфоглюконат
СН2ОН
С
О
Н
С
ОН
Н
С
ОН
6-Фосфоглюконатдегидрогеназа
НАДФ+
НАДФН
+ Н+
СО2
Н2С
ОРО3Н2
Рибулозо- 5-ф
36.
Пентозофосфатный путь окисления глюкозыЗНАЧЕНИЕ
1. Источник электронов и протонов (энергия) в процессе тканевого
дыхания.
2. Наиболее активно реакции пентозофосфатного пути идут в цитозоле
клеток печени, жировой ткани, эритроцитах, коре надпочечников,
молочной железе при лактации, в гораздо меньшей степени в скелетных
мышцах.
3. Этот путь окисления глюкозы не связан с образованием энергии, а
обеспечивает анаболизм клеток. В связи с этим у новорожденных и
детей первых лет жизни его активность довольно высока.
37.
Реакции окислительного пути протекают тольков том случае, если восстановленный кофермент
НАДФН возвращается в исходное окисленное
состояние НАДФ+ при участии НАДФН зaвиcимыx дегидрогеназ (т. е. при условии
использования гидрированного НАДФН в
восстановительных процессах). Если
потребности клетки в НАДФН незначительны,
рибозо-5-фосфат образуется в результате
обратимых реакций неокислительного этапа
пентозофосфатного пути, используя в качестве
исходных веществ метаболиты гликолиза —
глицеральдегид-3-фосфат и фруктозо-6-фосфат.
38.
Пентозофосфатный путь окисленияглюкозы
А. ОКИСЛИТЕЛЬНЫЙ ЭТАП
Б. НЕОКИСЛИТЕЛЬНЫЙ ЭТАП
(вместе А и Б – циклический процесс – пентозофосфатный цикл, где за 1
цикл полностью распадается 1 моль глюкозы)
6Глю-6-ф+ 12НАДФ+ +7Н2О
5 глю-6-ф+12НАДФНН+ +6СО2 +Фн
39.
Баланс аэробного гликолиза1. Аэробный гликолиз – субстратное
глю
2 пир
фосфорилирование
2. 2 пир
2 CH3COSKoA – окислительное
декарбоксилирование
3. Регенерация 2НАД·Н + Н+ в челночных
механизмах
2АТФ
6АТФ
36-38
АТФ
6-8АТФ
CO2
4.ЦТК
2 CH3COSKoA
24АТФ
H2O