































































































 biology
biologySimilar presentations:










Физиология возбудимых тканей
1.
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙДоцент кафедры нормальной
физиологии ФГБОУ ВО УГМУ
Минздрава РФ
к.м.н. Гагарина Е.М.
2.
Введение в курс физиологии• Нормальная физиология –
это наука о процессах
жизнедеятельности
целостного организма и
механизмах ее регуляции
для оптимального
приспособления к
меняющимся условиям
окружающей среды
3.
ВведениеРазделы нормальной физиологии
1. Общая физиология (физиология клетки,
эволюционная физиология, возрастная
физиология)
2. Частная физиология ( физиология крови,
физиология сердца, и др.)
3. Специальная физиология (физиология труда,
авиакосмическая физиология), т.е.
учитывающая род занятий и место проживания
4.
История физиологии• Уильям Гарвей (1578-1657) –английский врач,
анатом, физиолог и эмбриолог.
• Написал всемирно известную монографию
«Анатомические исследования о движении
сердца и крови» (1628 г)
Он писал: « Моим учением проложен путь более
одаренным, которые воспользуются им для
лучшего изучения вопроса»
5.
История• Иван
Михайлович
Сеченов (18291905) основоположник
отечественной
физиологии
6.
История• Иван Петрович
Павлов (1849 -1936)
-академик РАН,
лауреат
Нобелевской
премии, создатель
учения о высшей
нервной
деятельности
человека и
животных, один
из
основоположников
физиологии
пищеварения и
физиологии сердца.
7.
Физиологиявозбудимых
тканей
8.
Возбудимые ткани -1Физиологический покой, физиологическая
активность.
• Биологические системы (организмы, органы, ткани
и клетки) – могут находиться в двух основных
состояниях: в состоянии физиологического покоя и
активности.
• Физиологический покой – это состояние, при
котором живой организм не проявляет признаков
присущей ему деятельности при отсутствии
специальных раздражающих воздействий извне.
• При изменениях внешней или внутренней среды,
т.е. при действии раздражителей живые
организмы приходят в активное или деятельное
состояние
9.
• Раздражимость – это способность живых организмов, клеток,тканей и органов реагировать на внешнее воздействие
изменением своих физико-химических и физиологических
свойств
• Раздражимость проявляется в изменениях текущих значений
физиологических параметров, величина которых превышает их
сдвиги при покое.
• Раздражимость является универсальным проявлением
жизнедеятельности всех без исключения биологических систем.
• Раздражимостью обладают как животные, так и растения,
низшие и высшие их формы.
• Во всех проявлениях жизнедеятельности животных и
растительных организмов раздражимость играет
исключительную роль, так как со свойствами раздражимости
клеток и тканей связаны все проявления роста, размножения,
передвижения и т.д.
10.
• Отдельные клетки и ткани организма в ходеэволюции приспособились осуществлять быстрые
и точные ответы на действие раздражителей. Они
получили название возбудимых клеток и тканей.
• В организме животных и человека возбудимыми
тканями являются нервная, мышечная и
железистая.
11.
Возбудимые тканиВозбудимые ткани:
1. Нервная ткань (нейрон, нервное волокно,
синапс)
2. Мышечная ткань ( скелетные мышцы, сердечная
мышца, гладкие мышцы)
3. Железистая ткань (все виды желез внутренней
и внешней секреции)
12.
Возбудимые ткани• Другие ткани организма, обладая свойством
раздражимости, т.е. изменением обмена веществ
и энергии в ответ на раздражение, не относятся к
возбудимым, так как они не способны быстро
реагировать на раздражение.
13.
Возбудимые ткани• Возбудимость - это способность организма,
органа, ткани или клетки быстро отвечать на
раздражение специфической реакцией –
возбуждением (генерацией нервного импульса,
сокращением, секрецией и др).
14.
Возбудимые ткани• Мерой возбудимости живой структуры является
пороговая сила раздражителя, которая вызывает
видимую ответную реакцию.
• Возбудимость и порог раздражения находятся в
обратных соотношениях – чем выше порог, т.е. чем
большую силу нужно приложить, чтобы вызвать
ответную реакцию, тем ниже возбудимость живой
структуры. И наоборот – чем ниже порог, тем выше
возбудимость.
15.
РаздражителиПонятие о раздражителях.
• Для возбуждения большинства возбудимых
клеток необходимо наличие внешнего (по
отношению к этим клеткам) раздражителя
(исключение - возбудимые структуры,
обладающие автоматией)
• Раздражитель – это любой внешний фактор,
который при определенных условиях может
вызывать возбуждение возбудимой структуры
(клетки, органа).
16.
Раздражители классифицируют по несколькимпризнакам:
1) по модальности, т.е. по характеру энергии,
свойственной раздражителю, они
подразделяются на химические (кислоты,
щелочи), осмотические, тепловые, электрические,
световые, звуковые, словесные, биологические
(медиаторы, гормоны, микробы);
2) по адекватности, т.е. по соответствию
раздражителя воспринимающим раздражение
рецепторам, они
бывают адекватными и неадекватными.
17.
Раздражители• В физиологических опытах чаще всего используются
электрические раздражители, т.к. они физиологичны, т.е. не
повреждают ткань и напоминают нервные импульсы, и легко
дозируются по силе, времени, крутизне нарастания (т.е. скорости
нарастания силы тока во времени). Поэтому электрический ток
принято рассматривать как адекватный раздражитель.
18.
Электрические раздражители посиле бывают:
1) подпороговые (допороговые) раздражители – это слабые
раздражители, не вызывающие видимой ответной реакции. В ткани
возникает электротонический потенциал, а затем к нему присоединяется
локальный ответ (местное возбуждение)
2) пороговые раздражители – это минимальной силы раздражители,
вызывающие генерацию потенциала действия и минимальный
специфический физиологический эффект;
3) надпороговые раздражители – это более сильные раздражители,
вызывающие пропорциональное увеличение физиологического эффекта;
19.
Возбудимые тканиВозбуждение и его проявления.
• Если на возбудимую структуру (например, нейрон,
нервное волокно, мышечное волокно)
подействовать пороговым раздражителем, то
сформируется специфичный для данной структуры
тип ответной реакции.
• При этом возбудимая структура перейдет от
состояния относительного покоя к определенной
деятельности, т.е. к возбуждению.
20.
Возбудимые ткани• Возбуждение – это активный процесс, характеризующийся
сложным комплексом физико-химических и функциональных
изменений.
• Для каждой возбудимой структуры переход в возбужденное
состояние определяется осуществлением специфичной для
нее деятельности.
Например:
• мышца при возбуждении сокращается,
• эпителиальные, или железистые, клетки при возбуждении
выделяют секрет,
• нейроны генерируют нервный импульс, или потенциал
действия.
21.
Мембраны клеткиВсе возбудимые клетки покрыты снаружи мембраной ( ее
называют цитоплазматической или просто плазматической
мембраны, или плазмолеммой, или цитолеммой) .
• Внутри клетки также имеются мембранные структуры,
например, мембраны митохондрий, мембраны ядра клетки,
мембраны эндоплазматического ретикулюма. Однако по
своим свойствам они существенно отличаются от
плазматической мембран.
22.
Мембраны клеткиСтроение плазмолеммы
• Плазматическая мембрана всех
возбудимых клеток – самая толстая
из клеточных мембран. Ее толщина
составляет 7,5-11 нм.
• Под электронным микроскопом она
имеет вид трехслойной структуры,
представленной двумя электронноплотными слоями, которые
разделяются светлым слоем.
• Ее молекулярное строение
описывается жидкостно-мозаичной
моделью,
• Согласно этой модели, мембрана
состоит из двойного фосфолипидного
слоя, в который погружены и с
которым связаны молекулы белков.
23.
Мембраны клетки• Мембранные белки составляют более 40 % массы мембраны и
удерживаются в липидном бислое за счет гидрофобных
взаимодействий с молекулами липидов.
• По своему расположению относительно липидного бислоя
мембранные белки разделяются на две основные группы –
интегральные и периферические.
• Периферические белки находятся на поверхности мембраны и
непрочно связаны с ней.
24.
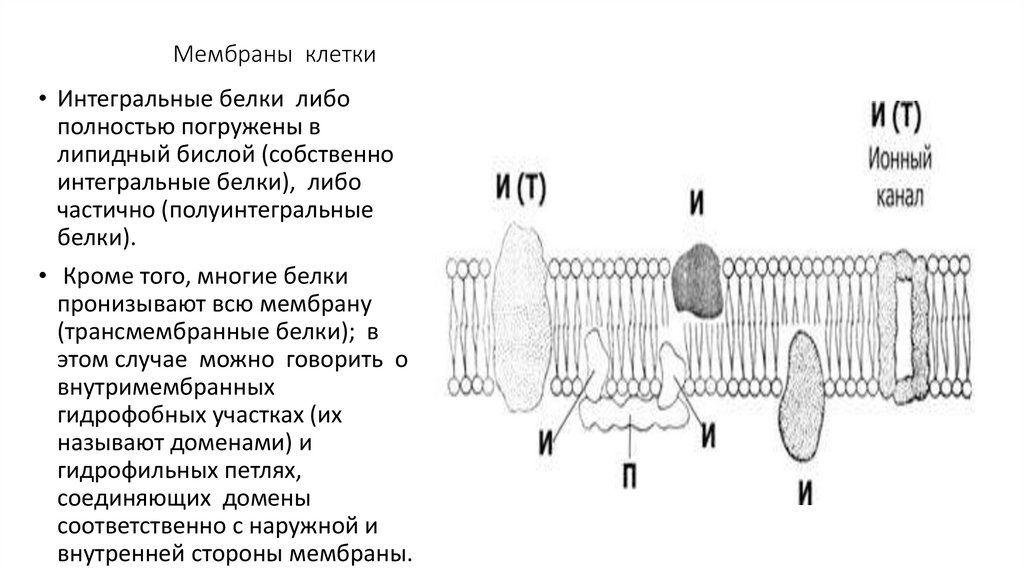
Мембраны клетки• Интегральные белки либо
полностью погружены в
липидный бислой (собственно
интегральные белки), либо
частично (полуинтегральные
белки).
• Кроме того, многие белки
пронизывают всю мембрану
(трансмембранные белки); в
этом случае можно говорить о
внутримембранных
гидрофобных участках (их
называют доменами) и
гидрофильных петлях,
соединяющих домены
соответственно с наружной и
внутренней стороны мембраны.
25.
Мембраны клетки• Часть мембранных белков (так называемые
гликопротеины) связана с молекулами
олигосахаридов, которые выступают за пределы
наружной поверхности плазмолеммы, а часть белков
(липопротеины) имеет липидные боковые цепи.
• Кроме того, молекулы олигосахаридов связаны с
липидами мембран, образуя гликолипиды.
• Выступая на наружную поверхность мембраны,
углеводные (олигосахаридные) участки
гликопротеинов и гликолипидов придают
поверхности клетки особое состояние, а также
обеспечивают формирование гликокаликса
(дословно – сладкая оболочка), который служит
своеобразным фильтром для поверхностной
мембраны, а также для рецепции химических
сигналов.
26.
Мембраны клеткиМембранные белки выполняют:
1) функцию ионных каналов, которые обеспечивают
пассивный транспорт ионов по градиенту
концентрации из наружной среды в клетку или
наоборот.
2) функцию активного транспорта ионов (ионные
насосы, например, Na/K- насос).
Т.е. белки выступают в роли пассивных или
активных переносчиков ионов и других
гидрофильных веществ, транспорт которых через
гидрофобные участки мембран запрещен
27.
Мембраны клетки3) функцию рецептора, т.е. специализированной
структуры, предназначенной для узнавания
определенных молекул (например, адреналина,
ацетилхолина), и передачи сообщения об этом
событии внутрь клетки к ее эффекторам,
например, к ферменту аденилатциклазе, которая
способствует образованию циклического 3,5аденозинмонофосфата (цАМФ) из
аденозинтрифосфата (АТФ).
4) функцию ферментов, осуществляющих перенос
определенных групп от одних молекул к другим
(например, перенос богатой энергией фосфатной
группы от АТФ к протеинкиназе).
28.
Мембраны клеткиОсновные функции плазматической мембраны
1) создание необходимой для деятельности данной
клетки микросреды внутри этой клетки. (барьернотранспортная функция)
2) формирование мембранного потенциала,
кратковременное изменение которого представляет
основной признак возбуждения (потенциал
действия).
3) рецепция внешних сигналов, в том числе
поступающих от других клеток через специальные
устройства (синапсы) или через кровь, лимфу или
ликвор. В роли таких сигналов выступают молекулы
медиаторов, гормонов, биологически активных
веществ
29.
Мембраны клетки• Таким образом, с участием мембраны
осуществляется межклеточное взаимодействие в
организме.
• Наряду со способностью возбудимых клеток
формировать мембранный потенциал и
потенциал действия, способность к
межклеточным взаимодействиям обеспечивает
все многообразие видов деятельности ЦНС
30.
Транспорт веществ черезмембрану
31.
транспорт• Мембранный транспорт веществ – это переход
вещества из внеклеточной среды во
внутриклеточную, либо, наоборот, из
внутриклеточной во внеклеточную среду.
• Кроме того, различают транспорт веществ из
цитоплазмы во внутриклеточные отсеки
(департаменты) или, наоборот, из отсеков в
цитоплазму (например, выход ионов Са2+ из
терминальных цистерн саркоплазматического
ретикулюма в саркоплазму, происходящий в
мышечном волокне).
32.
транспорт• Любой вид транспорта определяется свойствами
переносимого вещества – его способностью
растворяться в воде, его размерами, химическими
свойствами, а также градиентом (разницей)
концентрации между наружной и внутренней
поверхностью плазматической мембраны.
33.
транспортОбщие принципы, лежащие в основе транспорта
веществ через плазматические мембраны
1.Гидрофобные вещества хорошо проходимы через
плазматические мембраны. Поэтому их транспорт
определяется преимущественно наличием и
направленностью градиента концентрации –
вещество движется согласно законам
термодинамики из области его высокой
концентрации в область, где концентрация этого
вещества ниже.
34.
транспорт2. Гидрофильные вещества не могут свободно
проходить через плазматические мембраны, даже
если они имеют небольшие размеры. Для их
транспорта необходимы либо специальные частицы
– транспортеры, либо специальные механизмы, в
основе которых лежит изменение формы клетки.
35.
транспорт3. Если перенос вещества происходит с участием
транспортной частицы (переносчика), то в этом
случае возможно два варианта.
• перенос по градиенту концентрации. Такой вид
транспорта не требует для своей реализации в
данный момент времени затраты энергии (она
затрачивается ранее, при создании такого
градиента); поэтому его условно называют
пассивным транспортом.
• перенос вещества против градиента его
концентрации. В этом случае необходима затрата
свободной энергии (используется энергия,
которая освобождается при гидролизе АТФ, т.е. в
результате диссоциации этой молекулы на АДФ и
неорганический фосфат). Этот вид транспорта
получил название активный транспорт.
36.
транспорт4. Процесс транспорта веществ через
плазматическую мембрану может регулироваться,
поэтому проницаемость для конкретного вещества
– величина, изменяемая во времени.
• Особенно этот принцип важен в отношении
ионов натрия, калия, кальция и хлора – в
возбудимых клетках имеются специальные
механизмы регуляции проницаемости мембраны
для указанных ионов, позволяющих менять ее в
широких диапазонах, в том числе до полного
прекращения транспорта иона).
• При этом существуют два основных механизма
такой регуляции –
• А) за счет изменения уровня мембранного
потенциала (потенциалзависимый механизм)
• Б) за счет активации специфических клеточных
рецепторов (рецепторуправляемый механизм).
37.
транспортКлассификация механизмов транспорта веществ
через биологические мембраны.
• Различают прямой и опосредованный транспорт.
• Прямой транспорт осуществляется без участия
переносчиков и без затраты энергии. Он
осуществляется путем диффузии или фильтрации,
т.е. по типу пассивного транспорта. Примером
такого вида транспорта является перенос
кислорода как жирорастворимого вещества.
• Опосредованный транспорт во всех случаях
совершается с участием переносчика. При этом в
одних случаях этот вид транспорта идет без
затраты энергии (облегченная диффузия), а в
других – с затратой энергии (активный транспорт).
38.
транспорт• Активный транспорт может проткать по механизму
первично-активного и вторично-активного
транспортов.
• При первично-активном транспорте энергия
затрачивается на перенос данного вещества
(например, транспорт ионов натрия с помощью
натриевого насоса)
• при вторично-активном транспорте энергия
тратиться на перенос одного вещества (например,
натрия), в результате чего возникает возможность
переносить и другое вещество (например,
глюкозу)
39.
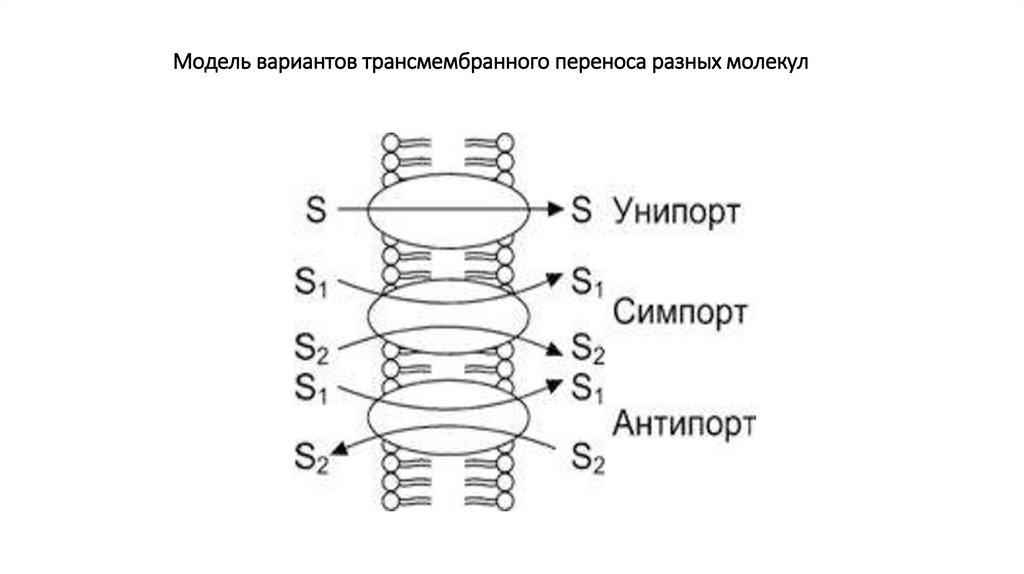
транспорт• При этом отдельно выделяют транспорт,
сопряженный с переносом двух веществ
(котранспорт), который может протекать по типу
симпорта (два вещества идут в одном
направлении, например, ионы Na+ с молекулой
глюкозы) или по типу антипорта (одно вещество
идет в клетку, второе – из клетки, например, так
работает Na/К-насос).
40.
Модель вариантов трансмембранного переноса разных молекул41.
транспорт• Вариант активного транспорта, т.е. транспорта с
затратой энергии, - это транспорт с изменением
архитектуры мембраны – экзоцитоз и эндоцитоз.
• В этом случае большая молекула, например,
белковая молекула, или группа молекул (квант
медиатора) выбрасывается за пределы клетки
(экзоцитоз; как пример – выброс медиатора из
пресинапса), либо поглощение этого вещества из
среды внутрь клетки (эндоцитоз; также
характерный для синапса).
42.
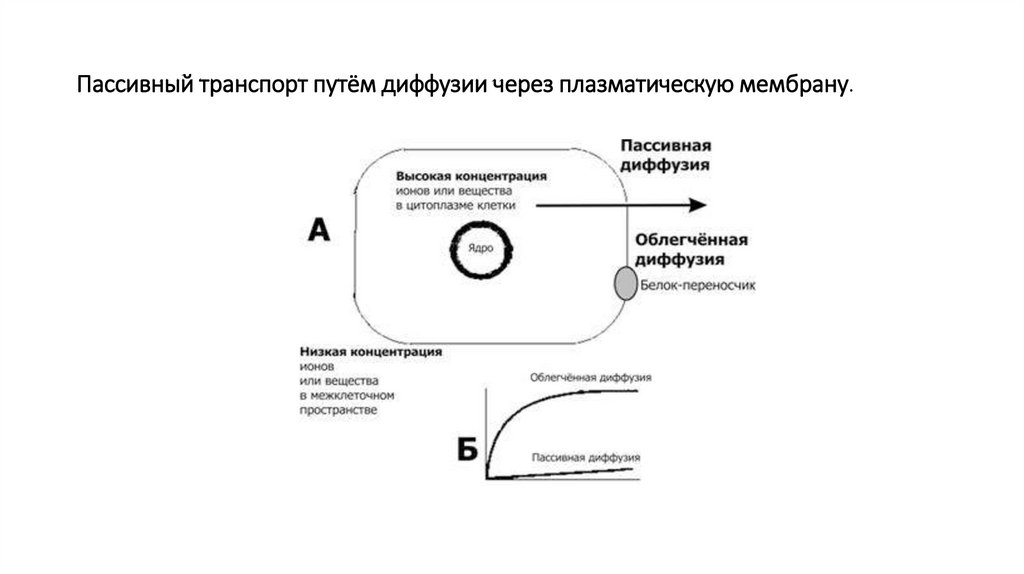
транспорт• Пассивный транспорт. Различают два его вида –
простую диффузию и облегченную диффузию.
• Механизмом простой диффузии осуществляется
перенос мелких жирорастворимых молекул (О2,
СО2, и других). Диффузия идет со скоростью,
пропорциональной градиенту концентрации или
градиенту напряжения транспортируемых веществ
(в соответствии с законом Фика).
43.
транспорт• Облегченная диффузия осуществляется через
специфические каналы (в том числе
специфические ионные каналы) или с участием
специфических белков-переносчиков. В том и
другом случае эти структуры являются
интегральными мембранными белками, а сам
перенос вещества идет без затраты энергии – за
счет химического или электрохимического
градиента.
• С помощью белков-переносчиков возбудимые
клетки (как и другие клетки) получают из
внеклеточной среды аминокислоты, моносахара
(например, глюкозу), которые транспортируются в
клетку по градиенту концентрации.
44.
Пассивный транспорт путём диффузии через плазматическую мембрану.45.
транспорт• Ионные каналы – это интегральные белки мембраны,
которые выполняют функцию транспортирующей
частицы для соответствующего иона.
• По сути, это тоже переносчики, но их способность
транспортировать ионы, т.е. ионная проницаемость,
регулируется с помощью специальных механизмов,
зависимых от уровня мембранного потенциала или от
состояния специфических клеточных рецепторов,
управляемых этими каналами.
• Изменение ионной проницаемости, особенно у
потенциалзависимых ионных каналов может
происходить с огромной скоростью – канал может
открываться только на 1- 3 мс.
46.
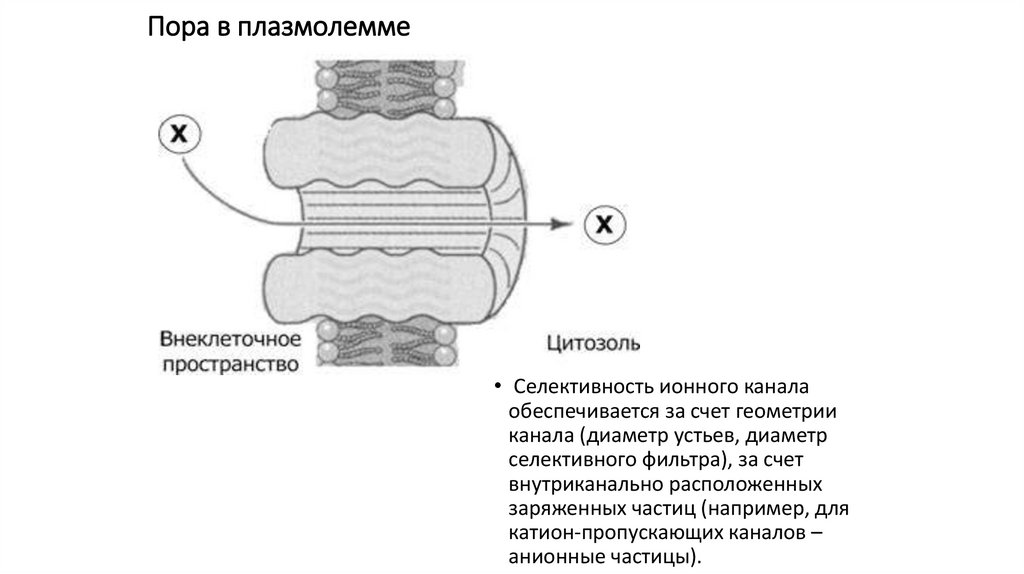
Пора в плазмолемме• Селективность ионного канала
обеспечивается за счет геометрии
канала (диаметр устьев, диаметр
селективного фильтра), за счет
внутриканально расположенных
заряженных частиц (например, для
катион-пропускающих каналов –
анионные частицы).
47.
транспорт• Каждый ионный канал состоит из собственно
транспортной системы и воротного механизма,
который открывает канал на некоторое время в
ответ на изменение мембранного потенциала,
или в ответ на связывание сигнальной молекулы
(лиганда) рецептором, либо на механическое
воздействие.
48.
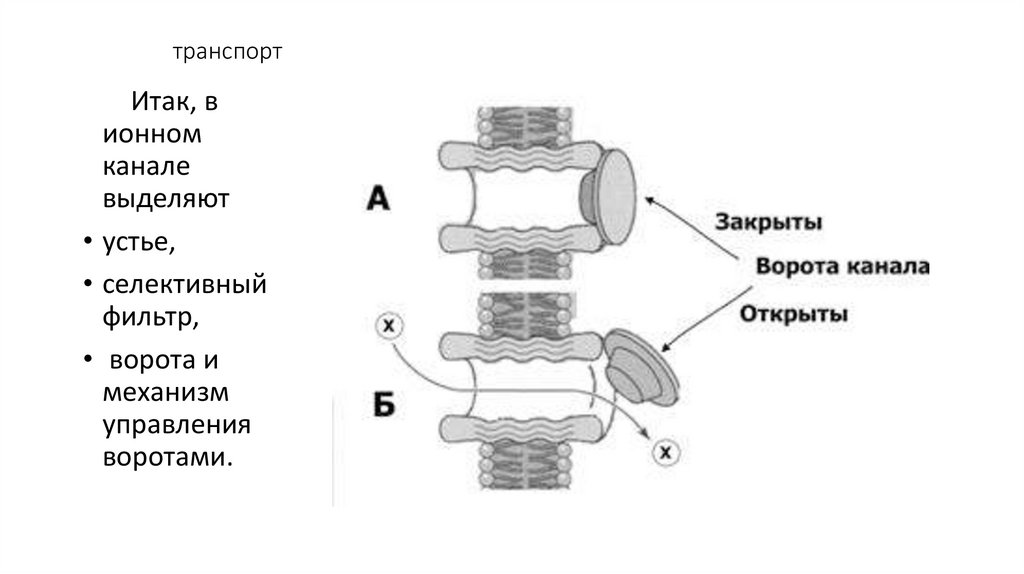
транспортИтак, в
ионном
канале
выделяют
• устье,
• селективный
фильтр,
• ворота и
механизм
управления
воротами.
49.
транспорт• Часть ионных каналов управляется за счет
разности потенциалов на мембране
(потенциалзависимые ионные каналы) – для этого
рядом с каналом имеется электрический сенсор,
который в зависимости от величины мембранного
потенциала либо открывает ворота каналов, либо
держит их закрытыми.
50.
транспорт• Второй вариант ионных каналов –
рецепторуправляемые каналы. В этом случае
ворота канала управляются за счет рецептора,
расположенного на поверхности мембраны: при
взаимодействии медиатора с этим рецептором
может происходить открытие ионных каналов.
51.
транспорт• Третий вариант ионных каналов –
механоуправляемые каналы, проницаемость
которых меняется под влиянием механической
силы (например, прилагаемой к микроворсинке
волосковой клетки внутреннего уха).
52.
Виды ионных каналов1. Электровозбудимые потенциалозависимые каналы
2. Хемовозбудимые
лиганд-рецептор-зависимые
каналы (рецепторуправляемые каналы)
3. Механовозбудимые, или механоуправляемые, каналы
53.
транспорт• Селективность ионных каналов во многом
определяется геометрией канала.
• Например, кристаллический диаметр иона натрия
– 0,19 нм, вместе с гидратной оболочкой его
диаметр достигает примерно 0,3 нм.
• Устье натриевого канала составляет 0,3-0,5 нм.
• Для того, чтобы пройти через такой канал
(главное – через его селективный фильтр), ион
натрия или другой ион должен освободиться от
гидратной оболочки и, только, в «голом» виде
может пройти через канал.
• Слишком «большой» ион не может войти в канал,
а слишком «маленький» – не способен отдать
гидратную оболочку в селективном фильтре,
поэтому не может выскочить из канала.
54.
транспорт• Направленность потока ионов определяется химическим и
электрохимическим градиентом.
• В частности известно, что в цитоплазме возбудимых клеток
концентрация ионов Na+ составляет 14 мМ, а во
внеклеточной среде – 140 мМ. Поэтому пассивный поток
ионов Na+ по натриевым каналам направлен из
внеклеточной среды в цитоплазму.
• Аналогичная ситуация для потока ионов Са2+, так как во
внеклеточной среде их концентрация намного больше (2,5
х10-3 М), чем в цитоплазме (10-7 М).
• Поток ионов К+ по калиевым каналам направлен,
наоборот, из клетки в среду, так как концентрация этих
ионов в цитоплазме намного больше, чем во внеклеточной
среде (150 мМ против 4-5 мМ)
55.
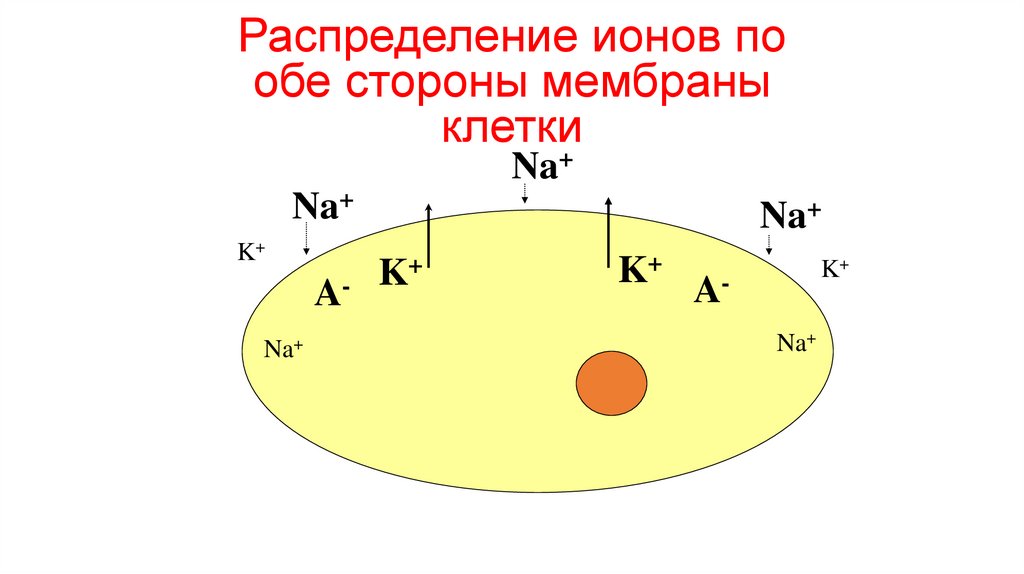
Распределение ионов пообе стороны мембраны
клетки
Na+
Na+
K+
ANa+
Na+
K+
K+
K+
ANa+
56.
Активный транспорт57.
активный транспортВиды активного транспорта
1) первично-активный транспорт, при котором
энергия гидролиза АТФ затрачивается на перенос
данного вещества против градиента его
концентрации,
2) вторично-активный транспорт, при котором
энергия на перенос данного вещества (например,
молекулы глюкозы) используется за счет
механизма переноса другого вещества (например,
натрия).
58.
активный транспорт• Первично-активный транспорт широко
представлен в организме - это:
• Na, К- насос многих плазматических мембран;
• Са-насосы плазматических мембран и мембран
саркоплазматического ретикулюма;
• натрий-водородный обменный механизм; натрийкальциевый обменный механизм;
• другие ионные насосы.
59.
активный транспорт• Все ионные насосы возбудимых клеток,
сконструированы и функционируют однотипно:
они представляют собой семейство белковферментов, которые получили общее название
ионтранспортирующих АТФаз Р-типа, или Е1-Е2типа.
• Эти насосы работают за счет энергии гидролиза
АТФ, которая затрачивается на изменение
конформационной структуры молекулыпереносчика, в результате которого меняется
сродство к переносимому иону и место
положения места связывания данного иона.
60.
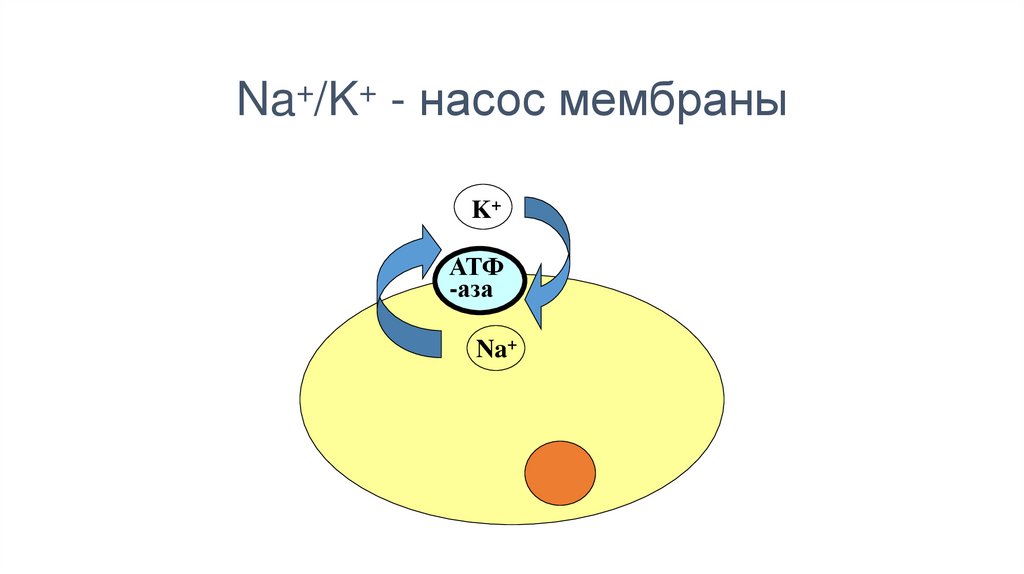
Na+/K+ –насос61.
Na+/K+ - насос мембраныK+
АТФ
-аза
Na+
62.
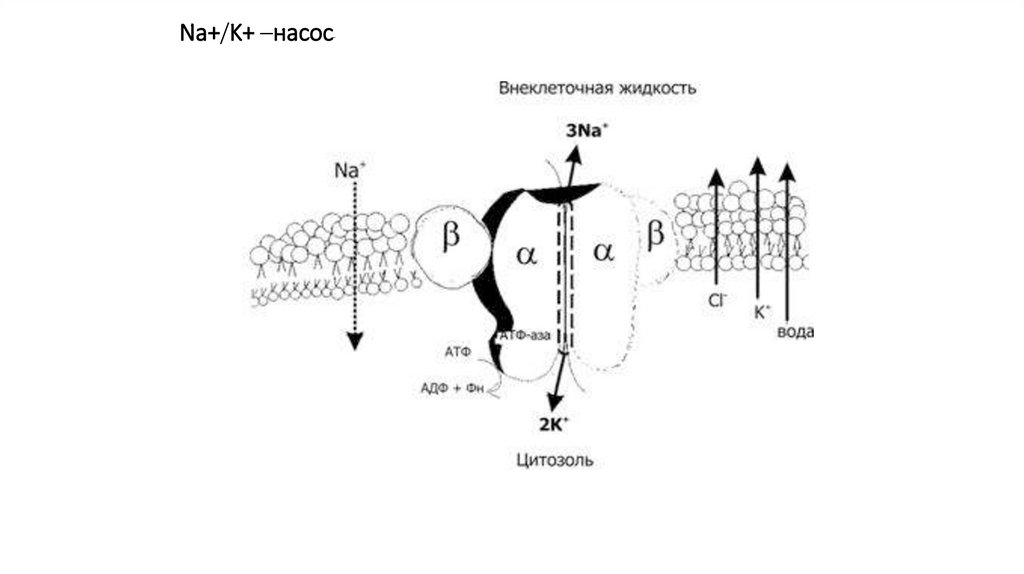
активный транспортРабота Na, К-насоса, или Na/К-АТФазы,
• Этот насос совершает активный перенос сразу
же двух ионов –
ионов Na+, которые переносятся против
градиента их концентрации из цитоплазмы в
окружающую среду,
• и ионов К+, которые, наоборот, переносятся из
наружной среды в клетку.
• Т.е этот насос «откачивает» ионы Na+ из клетки и
вкачивает в нее ионы К+.
63.
активный транспорт• Чтобы осуществить перенос натрия из клетки в среду, Na,
К- АТФ-аза внутри клетки, вследствие высокого сродства к
ионам Na+, присоединяет их к себе.
• В результате этого повышается АТФ-азная активность белкапереносчика, происходит гидролиз АТФ, это вызывает
высвобождение энергии, в результате меняется
конформационное состояние молекулы-насоса.
• Это приводит к транслокации мест связывания ионов Na+
на наружную поверхность клетки, снижению их сродства к
ионам Na+, уход их в окружающую среду.
64.
активный транспорт• Одновременно у молекулы-насоса появляется высокое
сродство к ионам К+ и к АТФ.
• Присоединение этих двух веществ к насосу вновь
вызывает конформационную перестройку, в результате
которой происходит повторная транслокация мест
связывания ионов К+ и снижение сродства к нему.
• Ионы К+ покидают насос внутри клетки, а места
связывания приобретают высокое сродство к ионам Na+.
• Цикл повторяется.
• Обычно Na, К-насос работает в электрогенном режиме –
на три выводимых из клетки иона Na+ внутрь клетки
вводится 2 иона К+.
65.
Электрогенез66.
История учения об электрогенезе• Луиджи Гальвани - «Трактат о силах электричества
при мышечном движении» (1791 г)
• Используя препарат из задних лапок лягушки,
соединенных позвоночником, он подвешивал его
на медном крючке к железным перилам балкона.
Когда лапки раскачивались ветром, то их мышцы
сокращались при каждом прикосновении к
перилам. Л. Гальвани заключил, что в спинном
мозгу лягушки формируется электрический ток,
приводящий к сокращению мускулатуры
( «балконный» опыт Л. Гальвани)
67.
История учения об электрогенезеАлександр Вольта (1792) доказал, что причиной
сокращения мышц в балконном опыте Л. Гальвани
является не биоэлектрический процесс, а
физическое явление - разность потенциалов
между разнородными металлами – медью
(крючок) и железом (перила балкона).
68.
История учения об электрогенезе• Позже Гальвани провел «истинный основной
опыт нервно-мышечной физиологии» (по Дю БуаРеймону Э.)
• Он стеклянной палочкой набрасывал нерв
препарата лягушки на слегка поврежденную
поверхность мышцы и наблюдал при этом ее
сокращение. («опыт без металлов») Это
доказывало наличие в живых клетках
электрических процессов (ток покоя, или ток
повреждения)
69.
История учения об электрогенезе• К. Маттеучи (1838) впервые показал, что наружная
поверхность мышечного волокна заряжена
электроположительно, а внутренняя –
электроотрицательно, т.е. показал наличие
мембранного потенциала или потенциала покоя
• В опыте «вторичное сокращение» он показал
существование тока действия (потенциал
действия). Он использовал два нервно-мышечных
препарата лягушки. Нерв первого препарата
помещался на раздражающие электроды, а нерв
второго препарата накладывался на мышцу
первого. В момент замыкания электрической цепи
наблюдалось сокращение мышц обеих лапок.
Электрический ток, вызывающий сокращение
мышцы второго препарата, был назван током
действия (потенциал действия) .
70.
История учения об электрогенезеСоздатели мембранной теории электрогенеза
(50-ые годы 20 века)
• Алан Ходжкин (Англия)
• Эндрю Хаксли (Англия)
• Бернард Катц (Англия)
• Джон Экклс (Австралия)
(Нобелевская премия 1963 года за разработку и
экспериментальное обоснование этой теории)
До настоящего времени создание мембранной
теории электрогенеза расценивается как
выдающееся достижение физиологии клетки и
физиологии возбудимых систем.
71.
Классификациябиопотенциалов
Мембранный потенциал (потенциал покоя)
Локальный (местный ) ответ
Потенциал действия
72.
Методы регистрациибиопотенциалов
73.
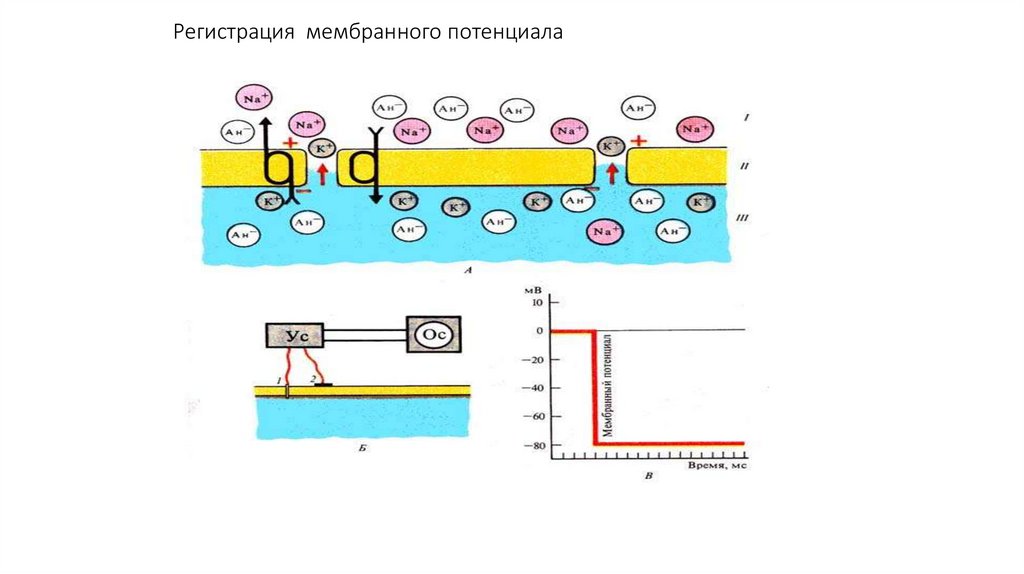
.Методы регистрации биопотенциалов
Методы регистрации биопотенциалов.
В связи с тем, что биопотенциалы относительно низкоамплитудны и
быстротечны, для их регистрации требуется
а) предварительно сигнал усилить
б) отразить его на приборе, позволяющем фиксировать быстро
протекающие процессы.
С этой целью в физиологии и клинической медицине широко
используются усилители биопотенциалов, осциллографы (катодные и
шлейфные), различные быстродействующие самописцы.
В последние годы в связи с созданием компьютерных технологий
возможности анализа биопотенциалов существенно возросли.
С 40-х годов 20 века в физиологии и клинической медицине
используются два основных метода регистрации биопотенциалов:
А) внеклеточный
Б) внутриклеточный.
74.
Регистрация мембранного потенциала75.
Мембранныйпотенциал
(потенциал покоя)
и его природа
76.
Мембранный потенциал и его природа• Мембранный потенциал, или потенциал покоя
(МП, ПП) - это разность потенциалов между
наружной и внутренней поверхностями мембраны
возбудимой клетки (нейроны, мышечные
волокна, кардиомиоциты, гладкомышечные
клетки, или миоциты, гландулоциты) в условиях
ее покоя
• Обычно у клеток возбудимых тканей МП
достигает -50-80 мВ (со знаком минус внутри
клетки).
77.
Потенциал покоя (Ео)мв
+30
0
-90
Ео
78.
Мембранный потенциал и его природа• Природа мембранного потенциала обусловлена
преимущественно ионами К+.,
• Это связано с тем, что в возбудимых клетках за
счет работы Na-К-насоса концентрация ионов К+ в
цитоплазме в условиях покоя поддерживается на
уровне 150 мМ, а во внеклеточной среде она
составляет 4-5 мМ., т.е. внутриклеточная
концентрация ионов К+ в 30-37 раз выше, чем
внеклеточная.
• Поэтому по градиенту концентрации ионы К+
стремятся выйти из клетки во внеклеточную среду
(в условиях покоя, действительно, существует
выходящий из клетки поток ионов К+, при этом
диффузия ионов происходит по калиевым
каналам, большая часть которых открыта)
79.
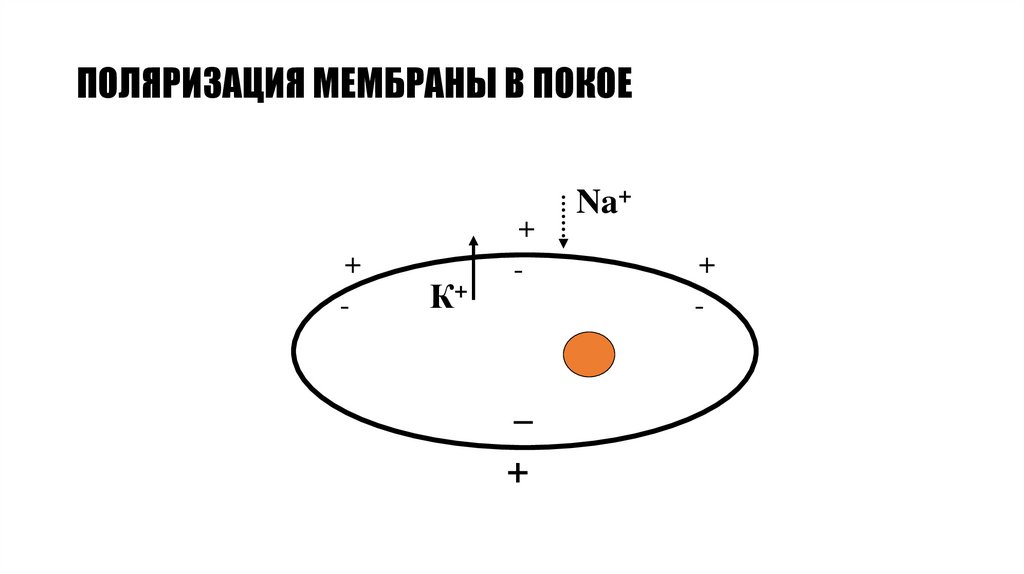
ПОЛЯРИЗАЦИЯ МЕМБРАНЫ В ПОКОЕ+
-
К+
+
-
_
+
Na+
+
-
80.
Мембранный потенциал и его природа• Так как мембрана возбудимых клеток непроницаема для
внутриклеточных анионов (глутамата, аспартата,
органических фосфатов), то на внутренней поверхности
мембраны клетки в следствие выхода ионов К+ образуется
избыток отрицательно заряженных частиц, а на наружной –
избыток положительно заряженных частиц.
• Возникает разность потенциалов, т.е. мембранный
потенциал, который препятствует чрезмерному выходу
ионов К+ из клетки.
• При некотором значении МП наступает равновесие
между выходом ионов К+ по концентрационному
градиенту и входом (возвратом) этих ионов по возникшему
электрическому градиенту.
• Мембранный потенциал, при котором достигается это
равновесие, получил название равновесного потенциала.
81.
Мембранный потенциал и его природа• Согласно мембранной теории возникновения
биопотенциалов, величина калиевого
равновесного потенциала (Ек), или собственно
величина мембранного потенциала может быть
рассчитана по уравнению Нернста:
• Ек = (RT: nF ) x ln ([Kе +] : [Ki +] ), где R – газовая
постоянная, T – абсолютная температура, F –
число Фарадея, n – валентность К+, равная 1, [Kе
+] – концентрация ионов К+ в среде, [Ki +] –
концентрация ионов К+ внутри клетки.
82.
Мембранный потенциал и его природа• После подстановки числовых значений констант и
перевода натурального логарифма в десятичный
эта формула имеет вид (для 37оС):
• Ек = - 61,5 log ([Kе +] : [Ki +] ).
• Т.е. величина равновесного калиевого потенциала,
а следовательно, и величина МП клетки
пропорциональна разности внеклеточной и
внутриклеточной концентрации ионов К+
• Чем эта разность больше, тем выше величина
мембранного потенциала.
83.
Калиевый равновесныйпотенциал мембраны по
Нернсту
[K+ out]
Е0 Ek 61,5 log --------- -90
mv
+
[K
in]
84.
Мембранный потенциал и его природаРоль ионов натрия и хлора в генезе мембранного
потенциала
Было замечено, что реальная величина мембранного
потенциала ниже расчетной (например, согласно расчетам,
величина МП спинальных нейронов лягушки должна быть
– 90 мВ, а реально она на 20 мВ ниже, т.е. составляет –70
мВ) .
• Согласно
уравнению
Нернста,
с
увеличением
концентрации ионов К+ во внеклеточной среде величина
мембранного
потенциала
должна
снижаться
пропорционально уменьшению градиента концентрации
этих ионов между клеткой и средой. Такая зависимость,
действительно, наблюдается, но при условии, что
концентрация ионов К+ в среде выше 20 мМ.
• Все это позволило утверждать, что помимо ионов К+
определенный вклад в создание мембранного потенциала
вносят ионы Na+ и Cl-
85.
Мембранный потенциал и его природаРоль натрия
• Концентрация ионов Na+ во внеклеточной среде
в 10 раз больше, чем внутри клетки (140 мМ
против 14 мМ).
• Поэтому ионы Na+ в условиях покоя стремятся
войти в клетку.
• Но основная часть натриевых каналов в условиях
покоя закрыта (относительная проницаемость
для ионов Na+ в 25 раз ниже, чем для ионов К+).
• Поэтому в клетку входит лишь небольшой поток
ионов Na+.
• Но этого достаточно, чтобы хотя бы частично
компенсировать избыток анионов внутри клетки.
86.
Мембранный потенциал и его природаРоль ионов хлора
• Концентрация анионов Cl- во внеклеточной среде
выше, чем внутри клетки (125 мМ против 9 мМ)
• Поэтому анионы Cl- стремятся войти в клетку,
очевидно, по хлорным каналам.
• Этот поток в условиях покоя относительно
небольшой (проницаемость для ионов Cl- в 2,5
раза ниже, чем для ионов К+),
• но за счет него вносится определенная часть
анионов и тем самым способствует росту
величины МП
87.
Мембранный потенциал и его природа• Таким образом, основными ионами, определяющими
величину МП, являются
• а) ионы К+, покидающие клетку.
• б) ионы Na+ , входящие в клетку в небольших
количествах, что частично уменьшает величину МП
• в) ионы Cl-,входящие в клетку в условиях покоя и в
определенной степени компенсируют влияние ионов Na+.
• Установлено, что чем выше проницаемость клеточной
мембраны для ионов Na+ в условиях покоя, тем ниже
величина МП.
• Именно этим обстоятельством объясняется сравнительно
низкий уровень МП у многих гладкомышечных клеток. Но
именно эта же причина позволяет гладкомышечным
клеткам спонтанно генерировать потенциал действия.
88.
Мембранный потенциал и его природа• Для того чтобы МП поддерживался на постоянном
уровне, необходимо поддержание ионного
гетерогенитета, т.е. ионной асимметрии.
• Для этого служат ионные насосы (Na-К-насос, а
также, вероятно, Cl-насос), которые
восстанавливают ионную асимметрию, особенно
после акта возбуждения.
• Так как этот вид транспорта ионов активный, т.е.
требующий затраты энергии, то на поддержание
мембранного потенциала клетки необходимо
постоянное наличие АТФ и механизма его
ресинтеза.
89.
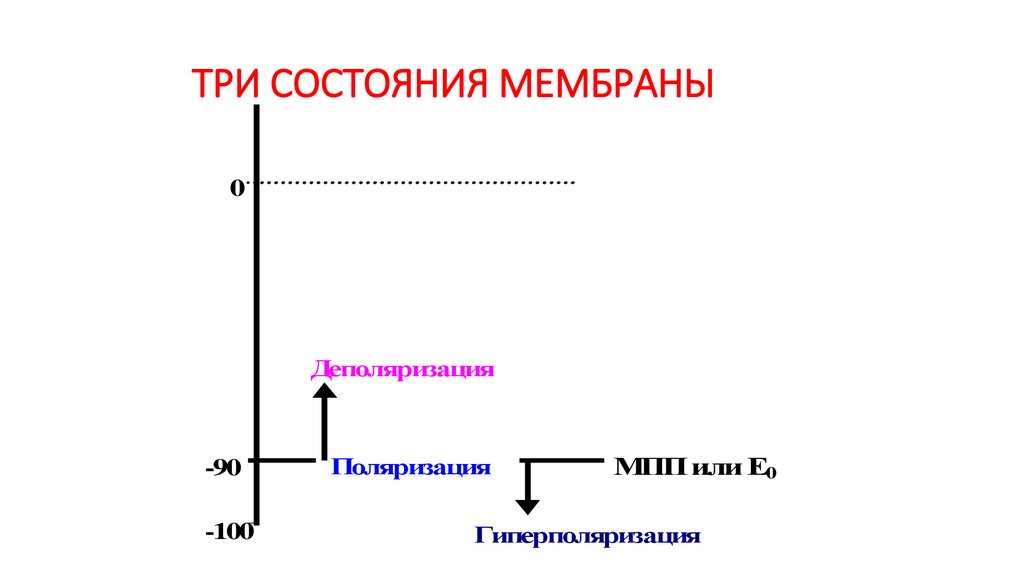
ТРИ СОСТОЯНИЯ МЕМБРАНЫ0
Деполяризация
-90
-100
Поляризация
МПП или Е0
Гиперполяризация
90.
Потенциал действия иего природа
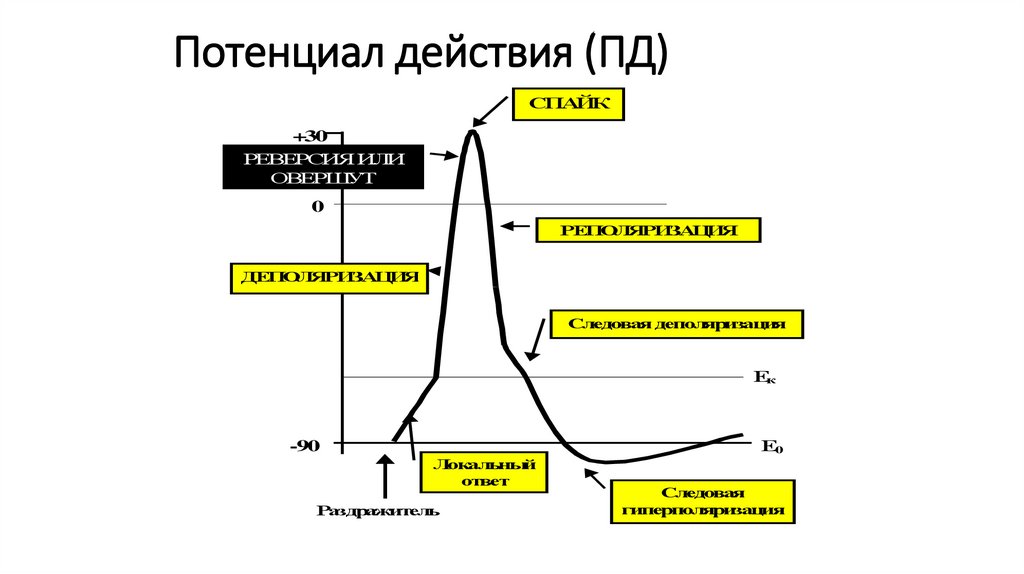
91.
Потенциал действияПотенциал действия (ПД).
• Потенциал действия - это кратковременное
изменение разности потенциалов между
наружной и внутренней поверхностями мембраны
(или между двумя точками ткани), возникающее в
момент возбуждения.
• Его форма и амплитудная характеристика
зависит от способа регистрации биопотенциала
92.
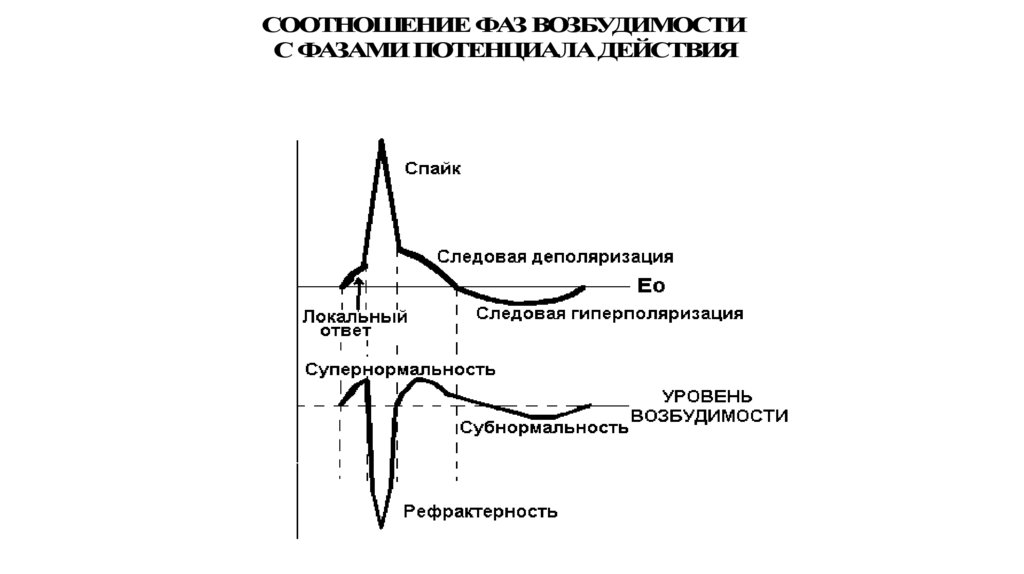
93.
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИС ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ
94.
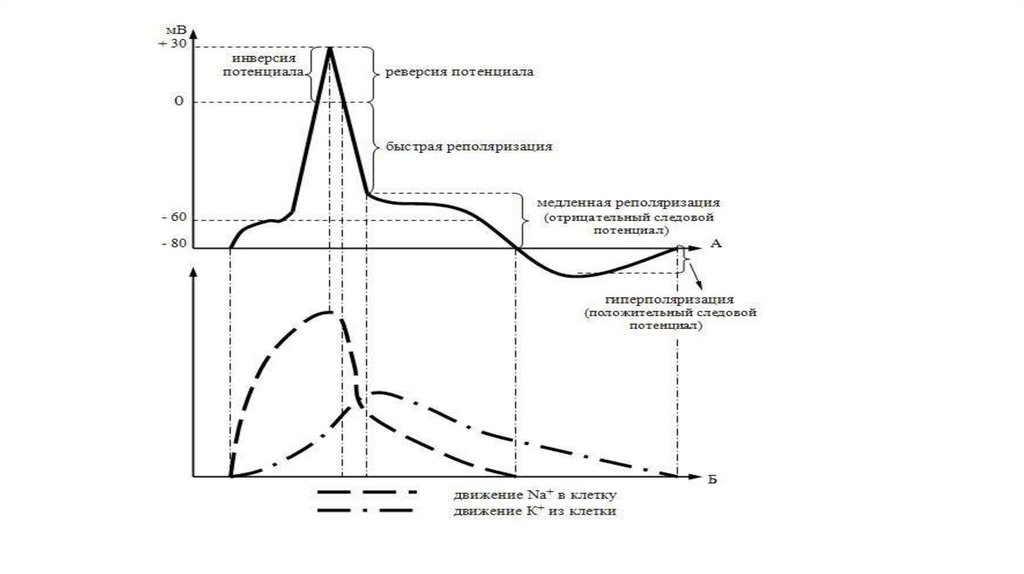
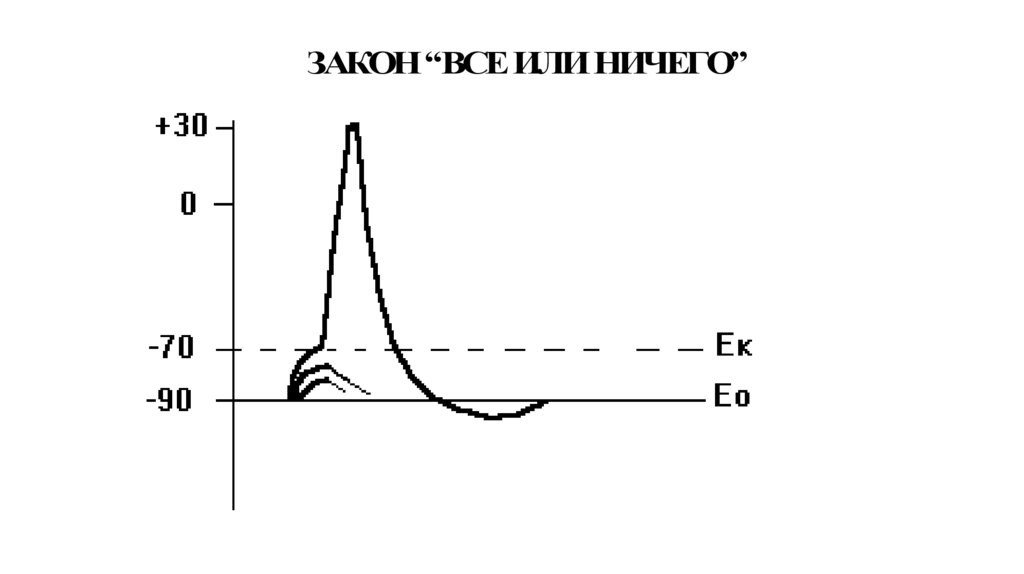
Потенциал действия (ПД)СПАЙК
+30
РЕВЕРСИЯ ИЛИ
ОВЕРШУТ
0
РЕПОЛЯРИЗАЦИЯ
ДЕПОЛЯРИЗАЦИЯ
Следовая деполяризация
Ек
Е0
-90
Локальный
ответ
Раздражитель
Следовая
гиперполяризация
95.
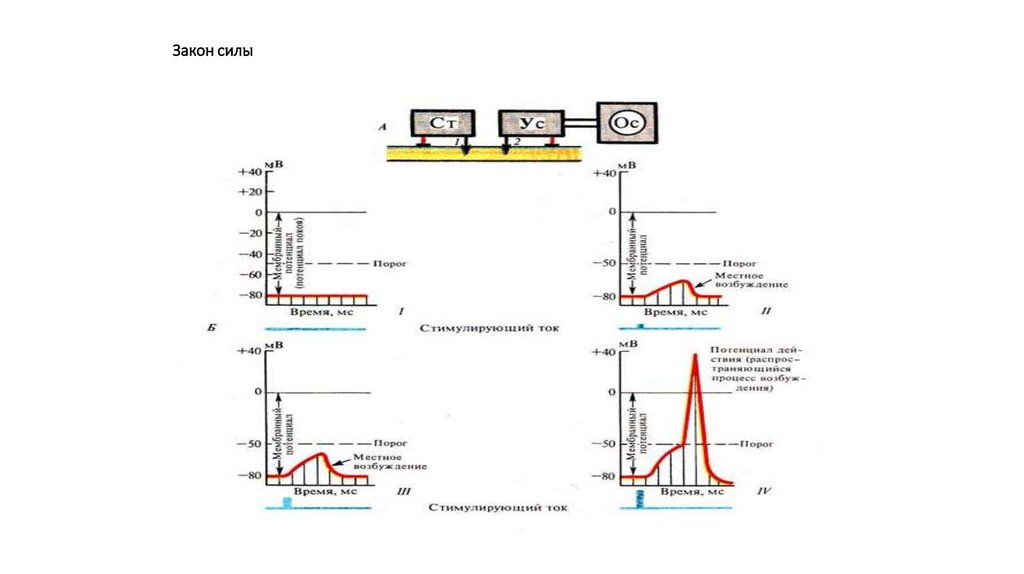
Потенциал действияВ пикообразном ПД выделяют
1. Локальный ответ – начальный этап деполяризации.
2. Фазу деполяризации – быстрое снижение мембранного
потенциала до нуля и перезарядка мембраны (реверсия,
или овершут)
3. Фазу реполяризации – восстановление исходного уровня
мембранного потенциала.
При этом в фазе реполяризации выделяют
а) фазу быстрой реполяризации ( вместе с фазой
деполяризации они составляют спайк ПД или пик ПД
б) фазу медленной реполяризации (следовые
потенциалы), в том числе следовую негативность
(следовую деполяризацию) и следовую позитивность
(следовую гиперполяризацию)
96.
Условия , необходимыедля генерации
потенциала действия
97.
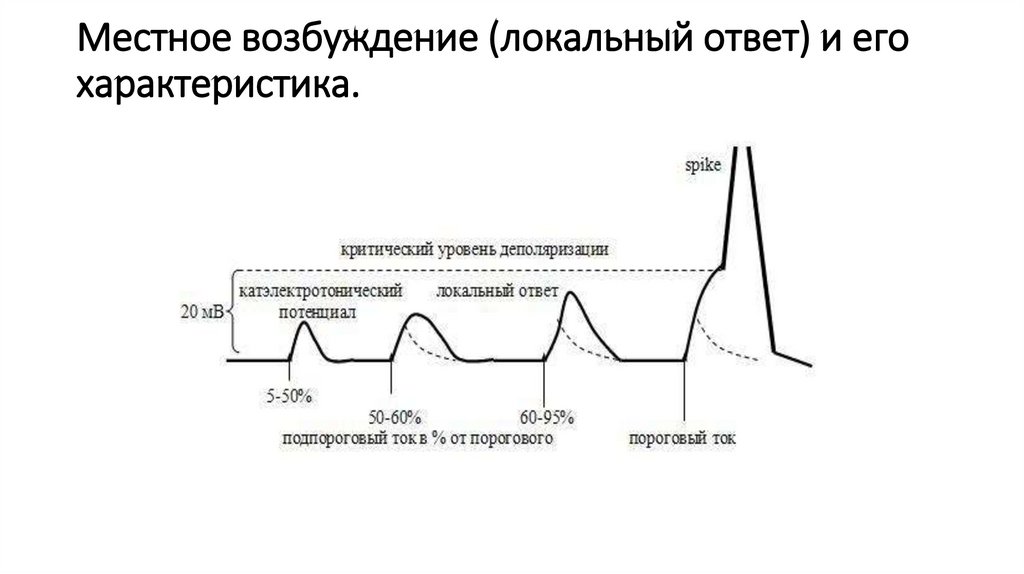
.Условия, необходимые для возбуждения
Условия, необходимые для возбуждения.
• Основным условием возбуждения возбудимой
клетки является снижение ее мембранного
потенциала до критического уровня
деполяризации (КУД).
• При достижении КУД возникающая в процессе
вхождения в клетку ионов Na+ и/или Са2+
деполяризация мембраны обязательно приведет к
тому, что все сенсоры, даже обладающие низкой
чувствительностью к величине деполяризации,
«сработают», в результате чего откроются все
ионные каналы, причастные к процессу
деполяризации во время возбуждении
98.
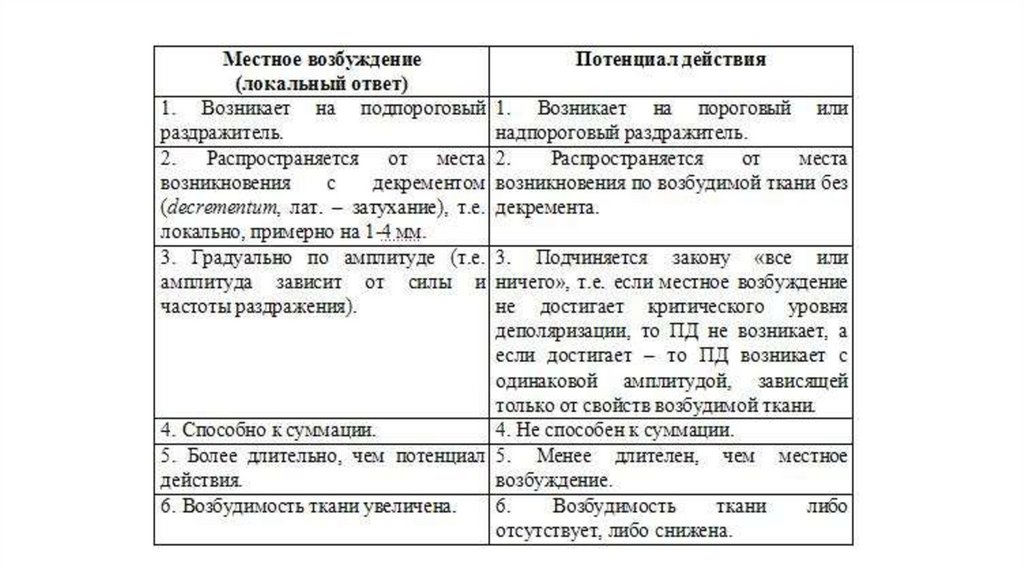
Местное возбуждение (локальный ответ) и егохарактеристика.
99.
100.
Изменение возбудимости ткани впроцессе её возбуждения
101.
Законы раздраженияПонятие о законах раздражения и их общая
формулировка.
Существует четыре основных закона раздражения
1) закон силы,
2) закон времени,
3) закон градиента
4) закон полярного действия тока (Пфлюгера).
• Все они отвечают на вопрос - каким должен быть
раздражитель, что бы он вызвал возбуждение ?
102.
Законы раздраженияОбъединенная формулировка закона силы,
времени и градиента гласит:
для того, чтобы раздражитель вызвал
возбуждение, он должен быть:
1) достаточно сильным (закон силы),
2) достаточно длительным (закон времени),
3) достаточно быстро нарастать (закон градиента).
Если эти условия не соблюдаются, то возбуждения
не происходит.
103.
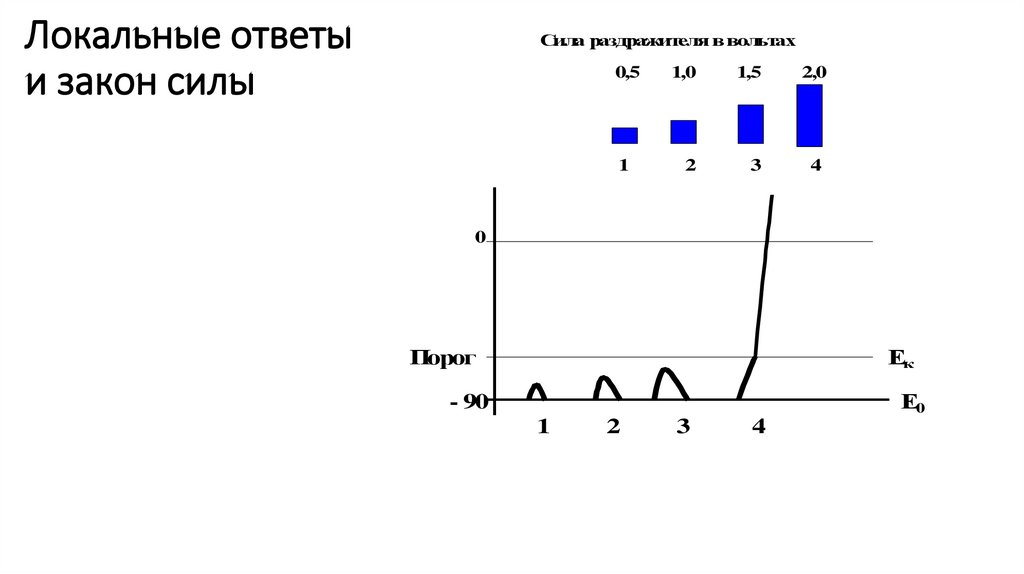
Закон силыЗакон силы.
Чтобы возникло возбуждение, раздражитель должен
быть достаточно сильным – пороговым или выше
порогового.
«Порог» - это минимальная сила раздражителя, которая
способна вызвать возбуждение.
Например, чтобы вызвать возбуждение нейрона при МП
= -70 мВ и КУД = -50 мВ, пороговая сила раздражителя
должна быть равной -20 мВ.
• Закон силы отражает основное условие, необходимое
для генерации потенциала действия – достижение
критического уровня деполяризации.
104.
Локальные ответыи закон силы
Сила раздражителя в вольтах
0,5
1
1,0
1,5
2
3
2,0
4
0
Порог
Ек
Е0
- 90
1
2
3
4
105.
Закон силы106.
ЗАКОН “ВСЕ ИЛИ НИЧЕГО”107.
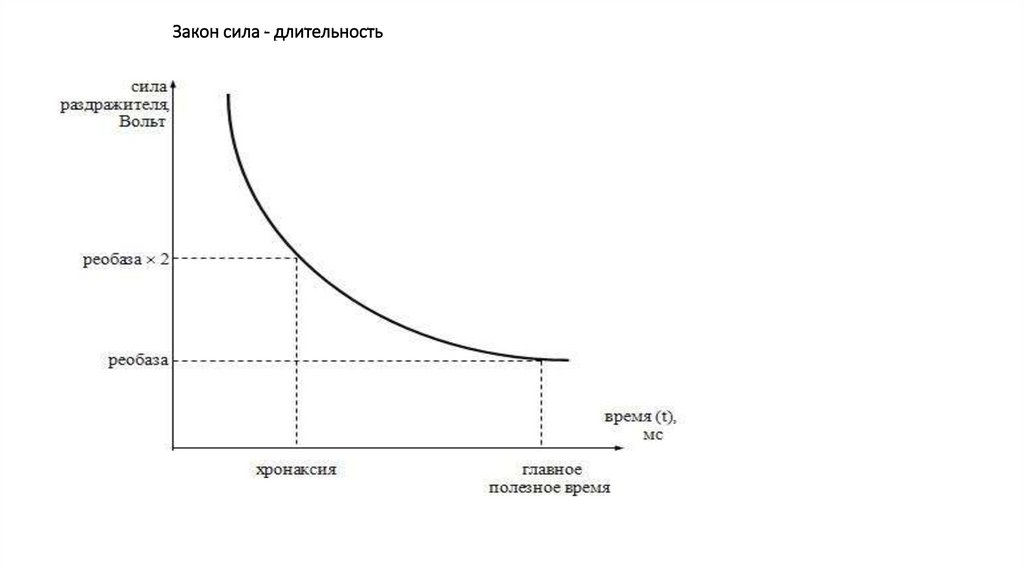
Закон времениЗакон времени, или зависимость пороговой силы раздражителя от времени
его действия.
• Раздражитель, вызывающий возбуждение, должен быть достаточно
длительным, т.е. для того, чтобы вызвать возбуждение, он должен
воздействовать на ткань не меньше некоторого минимального времени.
• В определенном диапазоне пороговая сила раздражителя находится в
обратной (гиперболической) зависимости от длительности его действия – чем
меньше по времени действует на ткань раздражитель, тем выше должна быть
его пороговая сила, необходимая для инициации возбуждения (зависимость,
или кривая Гоорвега-Вейса-Лапика)
• Кривая представляет собой отрезок гиперболы (ось абсцисс отражает
длительность стимула, а ось ординат –пороговую силу раздражителя,
вызывающего возбуждение).
108.
Закон сила - длительность109.
Закон времени. ХронаксиметрияХронаксиметрия.
• В клинической медицине и в физиологии реобаза и
хронаксия широко применяются для оценки возбудимости
тканей, например, при лечении ранений нервов.
• Для этих целей используется специальный прибор –
хронаксиметр, который позволяет определить
1) хронаксию и реобазу скелетных мышц (двигательная
хронаксия),
2) чувствительных нервных волокон (чувствительная
хронаксия),
3) вестибулярного аппарата (при раздражении
сосцевидного отростка),
4) сетчатки (вспышки света при ее электрической
стимуляции).
110.
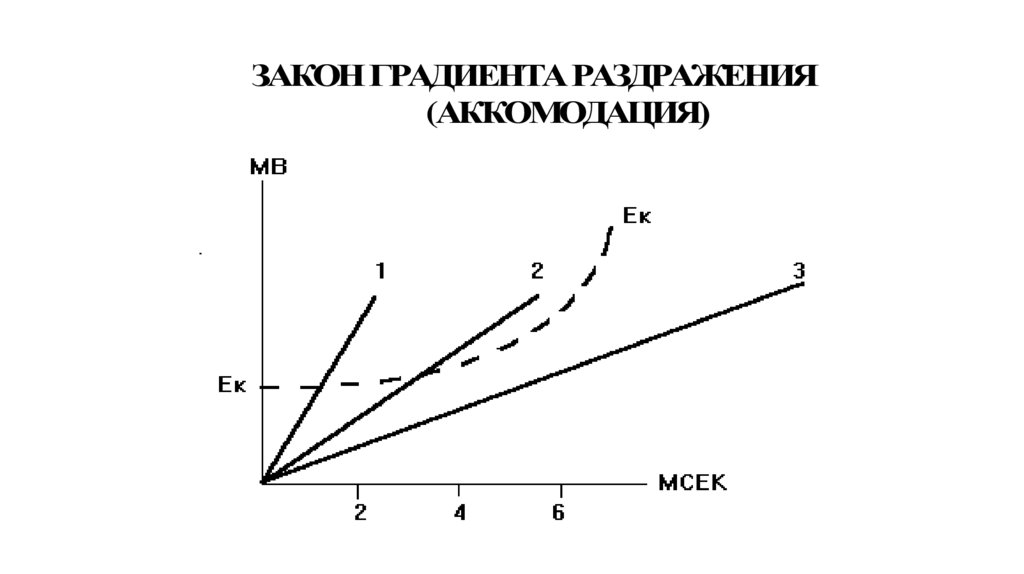
Закон градиентаЗакон градиента (аккомодации)
• Для того, чтобы раздражитель вызвал возбуждение, он
должен нарастать (по силе) достаточно быстро.
• Если раздражитель нарастает медленно, то из-за развития
аккомодации, т.е. инактивации натриевых каналов, порог
раздражения возрастает, поэтому для инициации
возбуждения величина стимула должна быть больше, чем
если бы он нарастал мгновенно.
111.
ЗАКОН ГРАДИЕНТА РАЗДРАЖЕНИЯ(АККОМОДАЦИЯ)
112.
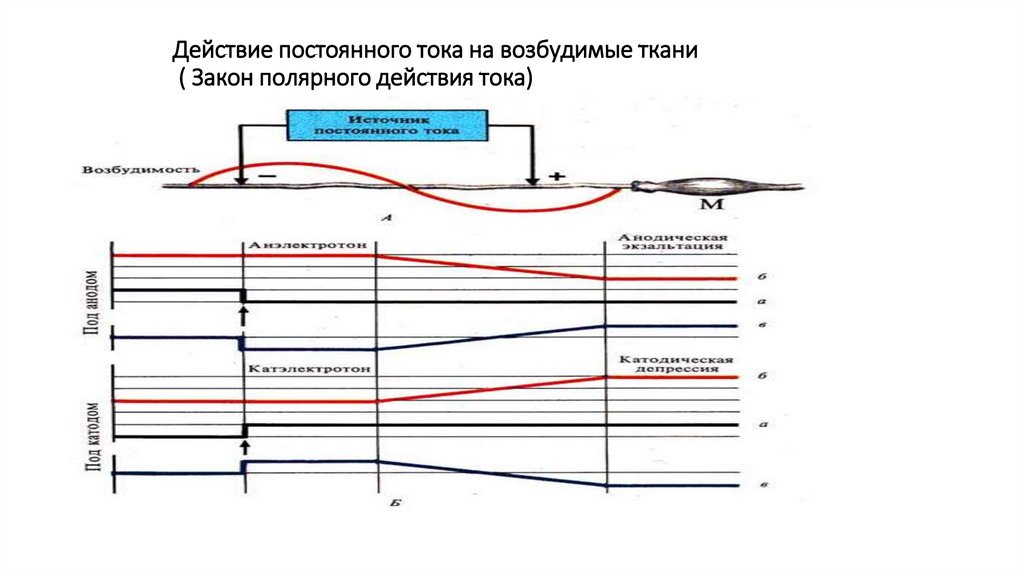
Действие постоянного тока на возбудимые ткани( Закон полярного действия тока)
Действие постоянного тока на возбудимые ткани.
• В 1859 г. Эдуард Ф.В. Пфлюгер сформулировал закон
полярного действия тока.
• Согласно этому закону, при действии постоянного тока
возбуждение происходит только в момент замыкания или
только в момент размыкания цепи постоянного тока. При
этом в момент замыкания оно возникает под катодом, а в
момент размыкания – под анодом.
• Этот закон позволяет объяснить механизмы развития
возбуждения. В частности, закон дает возможность понять
механизмы, лежащие в основе изменения КУД.
113.
Действие постоянного тока на возбудимые ткани( Закон полярного действия тока)
114.
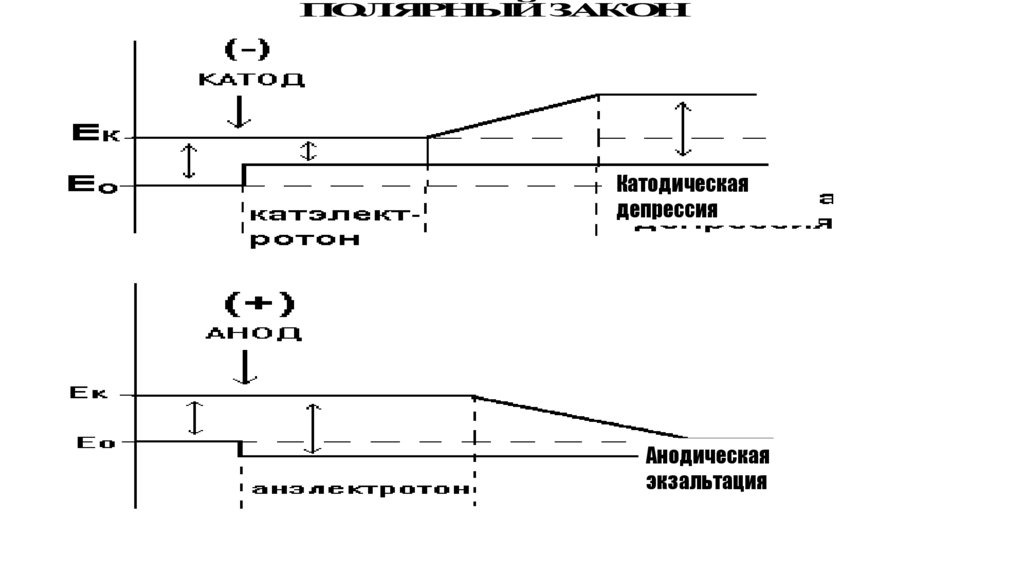
ПОЛЯРНЫЙ ЗАКОНКатодическая
депрессия
Анодическая
экзальтация
115.
Действие постоянного тока на возбудимые ткани.( Закон полярного действия тока)
• При замыкании цепи постоянного тока изменяется сила
раздражителя. Поэтому возбуждение возникает либо в
момент замыкания электрической цепи (под катодом),
либо в момент размыкания (под анодом). Это находится в
полном соответствии с законом градиента.
• При замыкании цепи постоянного тока под катодом
происходит деполяризация, и если она достаточна для
достижения КУД, то возникает возбуждение –
генерируется ПД.
116.
Действие постоянного тока на возбудимые ткани.( Закон полярного действия тока)
• Если возбуждение не происходит, то в области катода
изменяется возбудимость - в первые моменты
замыкания тока возбудимость под катодом вначале
возрастает (катэлектротон») . По мере прохождения
постоянного тока возбудимость под катодом резко
снижается (катодическая депрессия), что объясняется
процессом инактивации натриевых каналов, который
возникает в результате длительной деполяризации
мембраны в области катода.
• Явление катодической депрессии открыл пермский
физиолог Б.Ф. Вериго. Считается, что явление
катодической депрессии лежит в основе
пресинаптического торможения
117.
Действие постоянного тока на возбудимые ткани( Закон полярного действия тока)
• При замыкании цепи постоянного тока в области анода повышается величина
мембранного потенциала – возникает гиперполяризация.
• Поэтому в момент замыкании цепи возбуждение не возникает. Вследствие
гиперполяризации возбудимость под анодом снижается ( «анэлектротон»)
• Если постоянный ток действует сравнительно долго, то в условиях
гиперполяризации происходит снижение критического уровня деполяризации
(КУД), т.е. он приближается к исходному уровню мембранного потенциала. По
этой причине в момент размыкания цепи тока в области анода возникает
возбуждение, так как быстро уменьшающаяся величина мембранного
потенциала «достигает» КУД.
• Таким образом, явление анэлектротона и феномен появления возбуждения в
области анода при размыкании цепи постоянного тока вскрыли важный
механизм – изменение КУД в процессе функционирования возбудимых тканей.
118.
Действие постоянного тока на возбудимые ткани (Законполярного действия тока)
Практическое применение закона полярного действия
тока
• Если требуется заблокировать проведение возбуждения
по нерву (болевую рецепцию), то можно использовать
постоянный ток
• При этом в области расположения анода возбудимость
будет снижена, что приведет к блоку проведения
возбуждения.
• Аналогично – при длительной деполяризации в области
катода (когда возникает католическая депрессия) также
возбудимость будет снижена.
119.
Параметры возбудимости тканиДля оценки степени возбудимости тканей используют
следующие критерии:
1) порог раздражения;
2) хронаксия;
3) уровень лабильности (максимальное количество
импульсов, которое может воспроизвести ткань без
трансформации ритма);
4) скорость аккомодации.