






































 biology
biologySimilar presentations:










Генетика пола и сцепленных с полом признаков
1. Генетика пола и сцепленных с полом признаков
2. Хромосомная гипотеза наследственности
Сразу же после переоткрытия законовМенделя ученые обратили внимание на
факт удивительного сходства в поведении
менделеевских наследственных факторов
с закономерностями распределения
хромосом в процессе деления клетки. Уже
в 1902 году Уильям Сеттон и Теодор
Бовери, опираясь на этот факт,
обосновали хромосомную гипотезу
наследственности
3. Хромосомная теория наследственности
Основные экспериментальные доказательства того, что генынаходятся в хромосомах, были получены в исследованиях Т.Г.
Моргана и его учеников, выполненных на плодовой мушке
дрозофиле. Таким образом хромосомная гипотеза была
трансформирована в хромосомную теорию наследственности.
Т.Морган смог впервые объяснить результаты некоторых
несоответствий в расщеплении признаков в отдельных
вариантах скрещиваний дрозофил. Эти несоответствия
стандартному менделеевскому расщеплению оказались
характерны для т.н. признаков сцепленных с полом, т.е. для
признаков, определяемых генами, локализованными в
половых хромосомах.
4.
В биологии понятие «Пол» принятоопределять как
совокупность признаков и свойств
организма, обеспечивающих функцию
воспроизводства потомства и передачу
наследственной информации за счет
образования гамет
5. В онтогенезе пол особи может определяться:
а) до оплодотворения яйцеклетки сперматозоидом(прогамное) определение пола;
б) после оплодотворения (эпигамное) определение
пола;
в) в момент оплодотворения (сингамное)
определение пола.
6. Прогамно
До оплодотворения полопределяется у некоторых
организмов в результате
разделения яйцеклеток на быстро
и медленно растущие. Первые
(более крупные) после слияния с
мужской гаметой дают самок, а
вторые (мелкие) — самцов. У
коловраток, способных
размножаться помимо обычного
полового размножения с
оплодотворением,
партеногенетически, часть
партеногенетических яйцеклеток
во время развития лишается
половины хромосом. Из таких яиц
развиваются самцы, а остальная
часть дает начало самкам.
7. Эпигамно
У морского кольчатогочервя бонеллия
определение пола
происходит в процессе
онтогенеза: если
личинка садится на
дно, из нее развивается
самка, а если
прикрепляется к
хоботку взрослой
самки, то самец.
8.
Известно, что диплоидный набор хромосомвозникает в зиготе в результате слияния двух
гаплоидных гамет в процессе оплодотворения.
Гаплоидные наборы хромосом гамет оказываются
настолько соответствующими друг другу, что
каждая из хромосом зиготы оказывается в составе
пары (гомологичные хромосомы).
9.
Из этого следует принцип аллельного(парного) состояния гена у диплоидных
организмов, который не соблюдается
лишь для некоторых генов,
локализованных в половых хромосомах
при наличии т.н. гетерогаметного пола.
10. Гетерогаметность пола достигается цитологической и генетической неравноценностью пары половых хромосом
11. Половая дискриминация? Нет – компенсация дозы активных генов!
Чтобы сбалансироватьдействие генов Х хромосомы у
мужчин (где Х-хромосома
одна) и у женщин (где Ххромосомы две) природа
предусмотрела инактивацию
одной из Х-хромосом в каждой
клетке женского организма. У
женщин одна Х-хромосома
инактивирована, то есть гены
в ней не работают, а работают
только в другой хромосоме –
не инактивированной.
Инактивиация вызвана
компактизацией хромосомы
даже в интерфазе. Поэтому
компактизированная
хромосома видна при
окрашивании – в виде так
называемого тельца Барра
12. Типы хромосомного определения пола
Набор половыххромосом
Типичные представители
Самцы
Самки
ХY
ХХ
Млекопитающие,
большинство
рыб,
двукрылые
насекомые,
двудомные
растения.
ZZ
ZW
Птицы, бабочки, некоторые виды рыб,
земноводных, покрытосемянные растения.
Х0
ХХ
Большинство прямокрылых насекомых,
многие клопы (водяной клоп Protenor),
жуки, пауки, многоножки и нематоды.
ZZ
Z0
Отдельные
виды
насекомых (Fumea)
пресмыкающихся,
13. Гапло-диплоидия
Помимо этого унекоторых видов
насекомых (пчелы, осы,
муравьи) существует
особый т.н. гаплодиплоидный тип
определения пола, при
котором половые
хромосомы отсутствуют
вообще. У этих видов
при сперматогенезе
число хромосом не
редуцируется, т.о.
самки оказываются
диплоидны, а самцы –
гаплоидны.
Матка, рабочая пчела и
трутень
14. Функции половых хромосом
Помимо основной своей задачи определенияпола Х и У хромосомы выполняют и другие
функции. Природа экономна: кроме генов
влияющих на пол, в них расположены и
обычные гены. Признаки, развитие
которых определяют эти гены называются
сцепленными с полом.
15. «Гены находятся в хромосомах»
Изучение механизма наследованияпризнаков сцепленных с полом явилось
экспериментальным доказательством
хромосомной теории наследственности
16. Вернемся к опытам Т.Моргана, выполненным на дрозофиле
В Х-хромосоме этого вида известноболее 500 структурных генов. Для
самцов дрозофилы, имеющих в
норме половые хромосомы Х и У,
состояние большинства генов,
локализованных в Х-хромосоме
нельзя отнести ни к гомозиготному,
ни к гетерозиготному. Такое
состояние гена в Х-хромосоме у
организма с гетерогаметным полом,
при котором гены не имеют
аллельных гомологов, называется
гемизиготным. Наследование
гемизиготных генов происходит
своеобразно, в чем и убедился в свое
время Т.Морган (1909г.)
17.
В частности, Морган заметил, что наследованиеокраски глаз у дрозофилы зависит от пола
родительских особей, несущих альтернативные
аллели. Красная окраска глаз (доминантный ген
w+) доминирует над белой (рецессивный ген w).
При скрещивании белоглазых мух с мухами,
имеющими дикий тип по окраске глаз, были
получены различия в результатах реципрокных по
полу скрещиваний.
(Два скрещивания, различающиеся по тому, кто из родителей
вносит в зиготу доминантную или рецессивную аллель,
называются реципрокными).
18. Мутации глаз у Drozophila melanogaster
Мутации окраски иформы глаз у плодовой
мушки дрозофилы 1 дикий тип тускло
красные глаза (w+);
мутантные формы: 2 розовые глаза; 3 белые глаза (w);4 уменьшенные
«плосковидные» (Вar).
19. Так, при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки. Во
втором поколениинаблюдалось расщепление по фенотипу 3/1 (3 части красноглазых особей и 1
часть – белоглазых).
Р
Хw+ Хw+ x
гаметы P
Хw+
F1
Хw+ Хw х
гаметы F1 Хw+, Хw
F2
Хw+ Хw+, Хw+Хw,
красноглазая красноглазая
самка
самка
Хw У
Хw, У
Хw+У
Хw+, У
Хw+ У, Хw У
красноглазый
самец
белоглазый
самец
20. В реципрокном скрещивании красноглазого самца с белоглазой самкой в F1 наблюдалось расщепление 1:1, причем белыми глазами
обладали только самцы, акрасными – лишь самки. Т.о., в этом случае имело место т.н. крисс-кросс (или
крест-накрест) наследование, при котором сыновья наследуют признак матери,
а дочери - признак отца.
Р
Хw Хw
x
Хw+ У
гаметы P
Хw
Хw+, У
F1
Хw+Хw х
ХwУ
гаметы F1 Хw+, Хw
Хw, У
F2
Хw+ Хw, ХwХw, Хw+ У, Хw У
красноглазая белоглазая красноглазый
самка
самка
самец
1
1
1
белоглазый
самец
1
При дальнейшем скрещивании мух F1, между собой было получено поколение
F2, в котором с равной вероятностью встречались красноглазые и белоглазые
самцы и самки.
21.
Объяснить полученные результатынаблюдаемого расщепления по окраске
глаз Т.Морган смог, только
предположив, что ген, отвечающий за
окраску глаз, локализован в Ххромосоме, а Y-хромосома таких генов не
содержит.
22. Сцепленное с полом наследование при нерасхождении Х-хромосом
Продолжая эксперименты по скрещиваниюдрозофил, аспирант Т.Моргана Кальвин Бриджес в
1913г. обнаружил явление, которое нарушало схему
наследования «крест-накрест». С частотой 0,0010,1% в потомстве появлялись т.н.
«исключительные» мухи – красноглазые самцы и
белоглазые самки. Бриджес предположил, что во
время мейоза в направительное тело могут
отходить две Х-хромосомы, а в яйцеклетку – ни
одной. Второй возможный вариант возникает когда
в направительное тело Х-хромосомы не отходят, а
обе поступают в яйцеклетку. Это явление получило
название первичное нерасхождение хромосом
23. Сцепленное с полом наследование при нерасхождении Х-хромосом
РХw Хw
x
Хw+ У
Гаметы: самка/самец
Хw+
У
Хw
Хw+ Хw
красноглазая самка
Хw У
белоглазый самец
Хw Хw
Хw+Хw Хw
трисомия по Ххромосоме (гибнут)
Хw Хw У
белоглазая самка
-
Хw+
красноглазый самец
У
(гибнут)
24. Балансовая теория определения пола у дрозофилы
В 1921 году К. Бриджес обнаружил несколькофертильных самок, имевших триплоидный набор
хромосом (3X + 3A). В результате скрещивания
этих самок с нормальными самцами (2A + XY) в
потомстве среди нормальных самок и самцов были
обнаружены особи с промежуточным или
необычным проявлением половых признаков. Все
потомство распалось на восемь классов в
зависимости от соотношения половых хромосом и
аутосом.
25. Балансовая теория определения пола у дрозофилы
Р3X + 3A
х
ХУ + 2А
Гаметы:
самка/самец
Х +А
У+А
2Х +2А
3Х +3А
самка
2ХУ +3А
интерсекс
2Х + А
3Х +2А
сверхсамка
2ХУ + 2А
самка
Х +2А
2Х +3А
интерсекс
ХУ +3А
сверхсамец
Х+А
2Х + 2А
самка
ХУ + 2А
самец
26. К. Бриджес пришел к выводу, что пол определяется балансом числа половых хромосом и набором аутосом, а Y-хромосома у дрозофилы
вообще не играет роли в определении пола.27.
Балансовая теория оказаласьнеприменимой к человеку и высшим
растениям. У человека Y-хромосома
играет другую роль. Известно, что XX и
XY – соотношения половых хромосом у
человека приводят к образованию
нормальных женской и мужской особей
соответственно. Однако при отсутствии
Y-хромосомы формируется особь
женского пола при любом числе Xхромосом.
28. Синдром Шерешевского-Тернера
Особь X0 развиваетсяглавным образом по
женскому типу, но
имеет отличия от
нормы, известные под
названием «синдром
Тернера» (45,Х0).
Такие особи (девочки)
возникают с частотой
примерно 1/5000
новорожденных
29. Синдром Клайнфельтера
Синдром Клайнфельтеравстречается у одного из 400600 новорождённых
мальчиков и проявляется в
недоразвитии первичных и
вторичных половых
признаков, искажении
пропорций тела.
Клинические проявления
значительно варьируют:
большинство мужчин
47ХХУ внешне нормальны,
обладают нормальным
интеллектом, но
стерильны.
30. Гинандроморфизм
Еще одно доказательство того, что гены находятсяв хромосомах, было получено на мухах дрозофилах,
называемых латеральными (боковыми,
«сторонними») гинандроморфами. У таких мух
после первого деления яйцеклетки в одной из
клеток была утрачена Х-хромосома. Эта клетка
при последующих делениях дала клетки с
генотипом Х0, поэтому половина тела таких мух
сформирована как у самца и во ее всех клетках
цитологически наблюдается утрата одной из Ххромосом. Левая половина тела, происходящая из
клеток с нормальным генотипом ХХ, формируется
как самка.
31. Гинандроморфизм
В представленном на рисункеслучае муха гетерозиготна по гену
окраски глаз и по гену, влияющему
на форму крыла. Эти гены
расположены в Х-хромосоме.
Поэтому левый глаз мухи
неокрашен: проявилась
рецессивная мутация white,
присутствующая в одной из Ххромосом. Крыло изменено по
форме. Правый глаз окрашен и
крыло нормальной формы,
поскольку в нем остались обе Ххромосомы, в одной из которых
присутствует рецессивный ген
white, а в другой – доминантный ген
w+.
32. Признаки сцепленные с полом у человека
К настоящему времени в Х-хромосоме у человекаидентифицировано немногим более 1000
структурных генов, большинство из которых
детерминирует формирование нормальных и
патологических признаков. Генетический состав Ухромосомы значительно меньше - 78 генов, причем
54 из них имеют гомологию с генами Х-хромосомы.
Признаки, определяемые генами, находящимися в
Х и У- хромосомах сцеплены с полом и наследуются
отлично от аутосомных признаков.
33. Признаки сцепленные с полом у человека
В зависимости от природы гена и еголокализации можно выделить 3 основных
типа наследования признаков сцепленных
с полом у человека:
1. Х-сцепленный рецессивный тип
наследования;
2. Х-сцепленный доминантный тип
наследования;
3. Голандрический (У-сцепленный) тип
наследования
34. Х-сцепленный рецессивный тип наследования
Х-сцепленный рецессивный тип наследования характеризуется следующимипризнаками: 1) больные появляются не в каждом поколении;
2) больной ребенок рождается у здоровых родителей;
3) болеют преимущественно мужчины;
4) проявление признака (болезни) наблюдается преимущественно по
горизонтали;
5) вероятность наследования — у 25 % всех детей, в том числе у 50 %
мальчиков;
6) здоровые мужчины не передают болезни. Так наследуются у человека
гемофилия, дальтонизм, умственная отсталость с ломкой Х-хромосомой (с
Мартина-Белл), мышечная дистрофия Дюшенна, синдром Леша-Найхана и
др.
35. Дальтонизм
частичная цветовая слепота, один из видовнарушения цветового зрения. Д. впервые описан в
1794 Дж. Дальтоном, который сам страдал этим
недостатком. Д. встречается у 8% мужчин и у 0,5%
женщин. Предполагается, что в сетчатой оболочке
глаза существуют три элемента, каждый из
которых воспринимает только один из трёх
основных цветов (красный, зелёный, фиолетовый),
смешением которых получаются все
воспринимаемые нормальным глазом оттенки. Это
- нормальное, т. н. трихроматическое
цветоощущение. При выпадении одного из этих
элементов наступает частичная цветовая слепота
— дихромазия.
36. Гемофилия А
Гемофилия А - тяжелое заболевание, обусловленноедефектом фактора VIII свертывания крови.
Встречается с частотой 1:2500 новорожденных
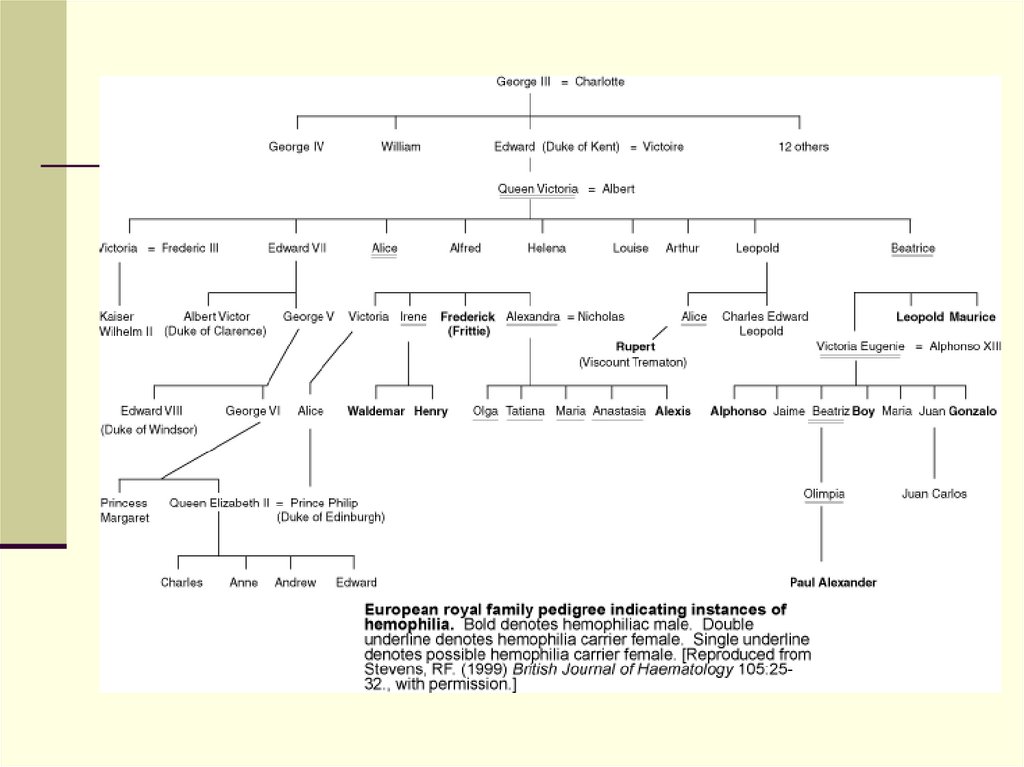
мальчиков. Ген расположен в длинном плече Ххромосомы (Xq28), структура его установлена.
Заболевание распознается обычно на 2-3-м году жизни.
Для него характерны множественные гематомы.
Преобладают кровоизлияния в крупные суставы
конечностей, подкожные, внутри- и межмышечные
гематомы, кровотечения при травмах и хирургических
вмешательствах, наличие крови в моче.
Кровоизлияния в полость суставов приводят к
развитию стойкой их тугоподвижности из-за
остеоартрозов (развитие соединительной ткани в
суставах).
37.
38. Механизм наследования гемофилии
39. Х-сцепленный доминантный тип наследования
Х-сцепленный доминантный тип наследования сходен саутосомно-доминантным, за исключением того, что мужчина
передает этот признак только дочерям (сыновья получают от
отца Y-хромосому). Примером такого заболевания является,
устойчивая к лечению витамином D особая форма рахита.
40. Голандрический тип наследования
Голандрический тип наследования характеризуется следующимипризнаками:
1) больные во всех поколениях;
2) болеют только мужчины;
3) у больного отца больны все его сыновья;
4) вероятность наследования у мальчиков 100 %. Так наследуются у
человека некоторые формы ихтиоза, гипертрихоз наружных
слуховых проходов и средних фаланг пальцев, некоторые формы
синдактилии (перепонки между пальцами ног) и др.