




































 biology
biologySimilar presentations:









")
Эукариоты и энергетический обмен
1.
БиологияЕГЭ
Урок 6
Эукариоты и энергетический обмен
2.
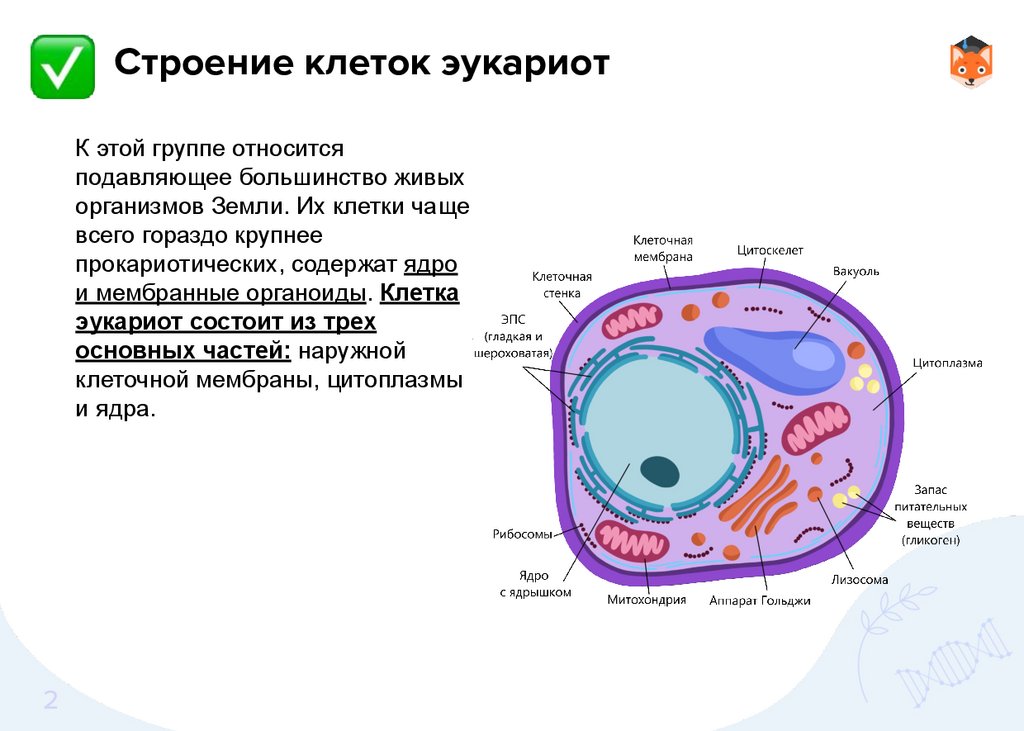
Строение клеток эукариотК этой группе относится
подавляющее большинство живых
организмов Земли. Их клетки чаще
всего гораздо крупнее
прокариотических, содержат ядро
и мембранные органоиды. Клетка
эукариот состоит из трех
основных частей: наружной
клеточной мембраны, цитоплазмы
и ядра.
2
3.
В клетке эукариот присутствуютследующие основные немембранные
органеллы:
➔
➔
➔
➔
3
рибосомы (молекулярные
машины для синтеза белка);
цитоскелетные нити;
построенные на основе
цитоскелетных нитей
структуры;
клеточный центр;
органоиды движения: жгутики,
реснички, миофибриллы
(сократимые нити поперечно полосатых мышечных
волокон).
4.
РибосомыРибосомы представляют собой мелкие органеллы,
состоящие из рибосомных РНК (рРНК) и рибосомных
белков. Они видимы в электронный, но не световой
микроскоп, т. к. имеют размеры порядка. Функция
рибосом — биосинтез белка. Работающая
рибосома включает в себя 2 субъединицы —
большую и малую. Они соединяются при синтезе
белка. До начала синтеза белка субъединицы
должны быть диссоциированы, то есть плавать в
цитоплазме отдельно друг от друга. Они могут быть
и соединены, но начать синтез белка могут, только
предварительно разъединившись.
Рибосомы про- и эукариот различаются по форме и
массе. Экспериментально это можно установить
путем центрифугирования.
4
5.
ЦитоскелетЭукариотическую клетку пронизывает система
структур, называемая цитоскелетом. Он
выполняет функции опоры, поддержания
формы клетки, движения (как всей клетки, так
и различных грузов внутри нее, в том числе
пузырьков и органелл).
Цитоскелет эукариотической клетки включает
белковые волокна 3 типов:
❏ микротрубочки;
❏ актиновые нити (тонкие филаменты);
❏ промежуточные филаменты.
5
6.
Все органеллы эукариотической клеткиможно условно разделить на три группы:
6
Одномембранные, стенка которых образована одной
мембраной. К ним относятся эндоплазматическая сеть,
аппарат Гольджи, лизосомы, вакуоли, секреторные пузырьки,
пероксисомы.
Двумембранные, стенка которых образована двумя
мембранами. Это митохондрии и пластиды (хлоропласты,
хромопласты и лейкопласты).
Немембранные. Это цитоскелет, клеточный центр, рибосомы
7.
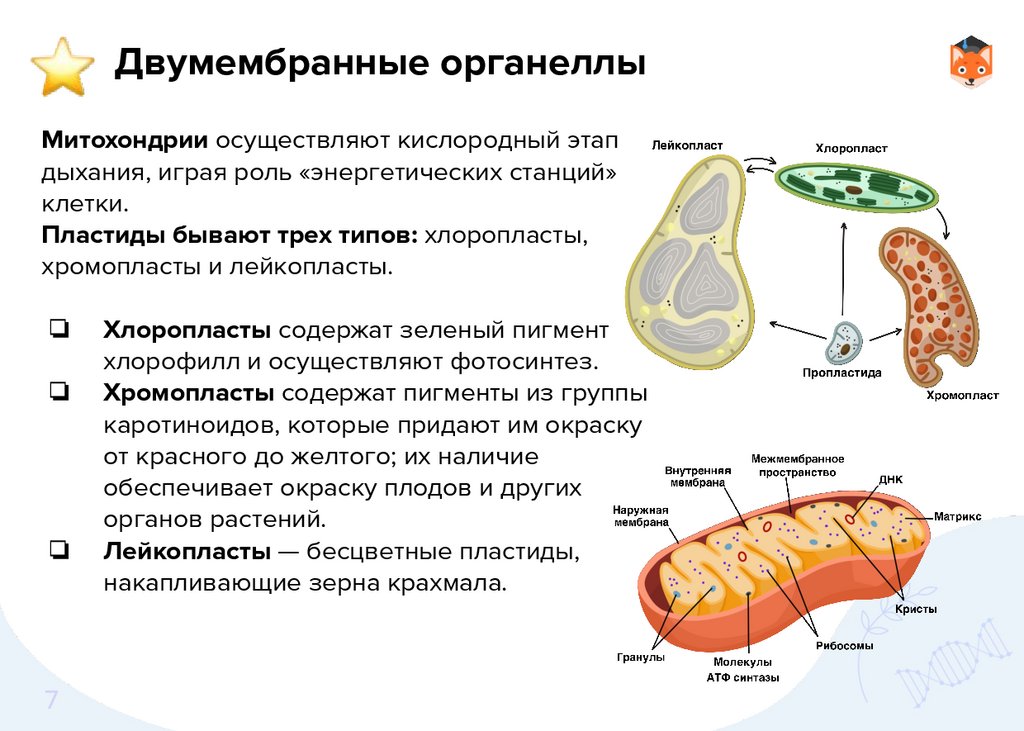
Двумембранные органеллыМитохондрии осуществляют кислородный этап
дыхания, играя роль «энергетических станций»
клетки.
Пластиды бывают трех типов: хлоропласты,
хромопласты и лейкопласты.
❏
❏
❏
7
Хлоропласты содержат зеленый пигмент
хлорофилл и осуществляют фотосинтез.
Хромопласты содержат пигменты из группы
каротиноидов, которые придают им окраску
от красного до желтого; их наличие
обеспечивает окраску плодов и других
органов растений.
Лейкопласты — бесцветные пластиды,
накапливающие зерна крахмала.
8.
Двумембранные органеллыДвумембранные органеллы называют также полуавтономными,
подчеркивая большую степень их самостоятельности в клетке.
Прежде всего, это означает, что они могут делиться. Новые
митохондрии и пластиды образуются только путем деления
существующих. У этих органелл имеется собственный геном —
кольцевая молекула ДНК, напоминающая ДНК бактерий. Она
содержит гены, кодирующие часть белков органеллы. Другая часть
кодируется в ядре и поступает из цитоплазмы (поэтому митохондрии и
пластиды не могут жить свободно, вне клетки).
Также эти органеллы имеют свой собственный аппарат синтеза
белка, то есть рибосомы. Эти рибосомы более мелкие, чем в
цитоплазме, и также похожи на рибосомы прокариот.
8
9.
Двумембранные органеллыВнешняя мембрана двумембранных органелл сходна
по составу с мембранами эукариот, внутренняя сходна с
мембранами прокариот.
Это согласуется с гипотезой о том, что внешняя
мембрана органеллы — это бывшая мембрана
пищеварительной вакуоли (фагосомы), где оказался
прокариотический симбионт, а внутренняя — это его
собственная мембрана.
Такой внутренний симбиоз называют эндосимбиозом, а
эту теорию происхождения двумембранных органелл —
теорией эндосимбиогенеза.
9
10.
МитохондрииСнаружи митохондрии покрыты внешней мембраной, сходной с
другими мембранами клетки.
Расположенная под ней внутренняя мембрана отличается по
липидному и белковому составу, напоминая мембрану бактерий. В ней
высоко содержание белков, именно в ней находятся ферменты,
осуществляющие окисление веществ кислородом, и синтез за счет
этого АТФ.
Для увеличения площади внутренняя мембрана митохондрий образует
многочисленные складки — кристы. Между двумя мембранами
находится изолированное от цитоплазмы межмембранное
пространство.
Внутри митохондрии находится пространство, называемое матрикс, в
котором осуществляется расщепление различных веществ: жирных
кислот, аминокислот и продуктов распада углеводов. В митохондриях
имеются собственные рибосомы и ДНК. Они ответственны за
образование части белков митохондрий.
10
11.
ПластидыПластиды имеют наружную и внутреннюю мембраны и
могут иметь внутренние мембранные мешочки
(тилакоиды и ламеллы), которые представляют собой
отпочковавшиеся внутрь впячивания внутренней
мембраны.
Все типы пластид образуются из пропластид, а также
могут превращаться друг в друга.
Система внутренних мембран хлоропласта включает в
себя совокупность мембранных каналов, называемых
ламеллами, и интенсивно окрашенные образования,
называемые гранами.
Граны представляют собой стопки плоских мембранных
пузырьков — тилакоидов, в мембранах которых
содержится хлорофилл и осуществляются
светозависимые реакции фотосинтеза. Внутренняя
полость хлоропласта называется строма. Она заполнена
раствором ферментов, осуществляющих темновые
(светонезависимые) реакции фотосинтеза.
11
12.
ЯдроЭто крупная органелла около 6–7 мкм диаметром. Оно окружено ядерной
оболочкой, которая образована двумя параллельно расположенными
мембранами.
Ядерная оболочка пронизана ядерными порами, где
мембраны смыкаются, и полость ядра сообщается с
цитоплазмой. В порах находятся сложные белковые
комплексы. Они переносят через оболочку из ядра в
цитоплазму крупные молекулы и молекулярные
комплексы,
такие как мРНК и рибосомы, а из цитоплазмы в ядро —
ядерные белки, которые синтезируются в цитоплазме.
Внутри ядра находится одно или несколько ядрышек— плотных
образований, где происходит синтез рибосомных РНК и сборка субъединиц
рибосом. Остальное пространство ядра
заполнено полужидкой кариоплазмой, в которой находятся молекулы
ДНК, соединенные со специфическими белками, — хроматин.
12
13.
ЯдроВ процессе клеточного деления нити хроматина укорачиваются и утолщаются,
превращаясь в хромосомы.
Перед делением хромосомы имеют Х-образную форму. Центральная часть, в которой
соединяются две половины хромосомы, носит название центромеры , или первичной
перетяжки. Кроме того, в хромосоме выделяются более плотные концевые участки,
называемые теломерами.
Видоспецифичный набор хромосом со всеми их характеристиками называется кариотипом.
Клетки могут содержать один набор хромосом или их кратное число. Число хромосомных
наборов называется плоидностью. Клетки, содержащие один набор хромосом, называются
гаплоидными, содержащие два набора — диплоидными, три набора — триплоидными,
четыре набора — тетраплоидными и т. д.
Чаще всего мы имеем дело с организмами, состоящими из диплоидных клеток: это
животные и большинство растений. Встречаются организмы, построенные из гаплоидных
клеток, например, мхи. Организмы, в клетках которых более двух наборов хромосом,
принято называть полиплоидами.
13
14.
Одномембранные органеллыОсновные структуры
эукариотической клетки:
цитоплазматическая мембрана,
цитоплазма и ядро.
Клетки эукариот содержат множество
внутренних структур, выполняющих
определенные функции. Эти структуры
называются органеллами.
14
15.
ЦитоплазмаЦитозоль (гиалоплазма) — свободная часть
цитоплазмы, в которой взвешены органеллы.
Цитоплазма — обязательная
часть клетки, заключенная между
плазматической мембраной и
ядром.
Цитоплазма объединяет все
клеточные структуры и
способствует их взаимодействию
друг с другом.
Цитоплазма заполнена взвесью
частиц и органелл.
15
16.
ЦитоплазмаПо химическому составу цитозоль представляет из себя
густой раствор белков, углеводов (глюкозы) и ионов,
заполняющий все внутреннее пространство клетки.
Концентрации ионов натрия и калия внутри клетки и во
внеклеточном пространстве различна, что играет важную
роль в осморегуляции и передаче сигнала.
У прокариот большинство реакций метаболизма протекает
в цитозоле, т. к. нет мембранных органоидов.
У эукариот часть химических реакций протекает в цитозоле, а
часть — внутри органоидов.
В животных клетках различают два слоя цитоплазмы:
★
эктоплазма — наружный слой цитоплазмы (мало
органоидов, высокая вязкость);
★
эндоплазма — внутренний слой цитоплазмы (содержит
основные органоиды)
16
17.
ЦитоплазмаЦиклоз — одно из основных свойств цитоплазмы живой клетки — способность
к движению, которое обеспечивает транспорт веществ и связь органоидов.
В цитоплазме осуществляются
все процессы клеточного
метаболизма, кроме синтеза
нуклеиновых кислот,
происходящего в ядре. Под
контролем ядра цитоплазма
способна к росту и
воспроизведению, при частичном
удалении она полностью
регенерирует. Цитоплазма, как
правило, не способна к
длительному автономному
существованию.
17
18.
Плазматическая мембранаСнаружи эукариотическая клетка, как и
прокариотическая, окружена цитоплазматической
мембраной. Она выполняет те же функции, что и у
прокариот: изолирующую, транспортную и рецепторную.
Рецепторная функция у эукариотических клеток развита
гораздо сильнее, чем у прокариот, поэтому в
цитоплазматической мембране у них гораздо больше
белков - рецепторов. У многоклеточных организмов
цитоплазматическая мембрана выполняет также
функцию межклеточного узнавания и
взаимодействия. У растений и грибов снаружи от
цитоплазматической мембраны лежит клеточная стенка.
У растений она построена на основе целлюлозы, а у грибов — на основе хитина. У животных
клеточной стенки нет, но к мембране снаружи прикрепляется довольно толстый слой специфических
полисахаридов и белков, называемый гликокаликс. В отличие от клеточной стенки, он эластичен, что
позволяет клеткам менять свою форму. В отличие от клеточной стенки, гликокаликс прочно связан с
мембраной и не отделяется от нее.
18
19.
Гликокаликс и его функцииГликокаликс — углеводная оболочка клетки. Большая часть углеводов
мембран представлена не свободными молекулами, а комплексами с
другими молекулами:
углевод + белок = гликопротеин (большая часть интегральных белков);
углевод + липид = гликолипид (1/10 часть липидных молекул мембраны);
белок с боковыми углеводными цепями = протеогликан.
Углеводные части мембранных структур почти всегда направлены
наружу и выступают над поверхностью клетки.
Функции гликокаликса:
● отталкивание от клетки отрицательно заряженных частиц (т. к.
многие углеводные молекулы гликокаликса имеют отрицательный
заряд);
● скрепление соседних клеток друг с другом;
● рецепторная функция (связывание гормонов, что приводит к
активации определенных мембранных белков, и запускание
каскада ферментативных реакций внутри клетки);
● участие в иммунных реакциях.
19
20.
Одномембранные органеллыЭти органеллы, как предполагается, в ходе
эволюции образовались путем впячивания
наружной мембраны внутрь и
отпочковывания этих впячиваний. Почти все
эти органеллы связаны между собой —
прежде всего системой пузырькового
(везикулярного) транспорта, когда пузырьки
отпочковываются от одной органеллы и
сливаются с другой, перенося содержимое и
компоненты мембраны. Все вместе эти
органеллы называются вакуолярной
системой (эндоплазматическая сеть, аппарат
Гольджи, лизосомы, вакуоли, секреторные
пузырьки, пероксисомы)
20
21.
Эндоплазматическая сетьОн представляет собой систему мембран, образующих соединенные между собой цистерны,
полость которых не сообщается с окружающей цитоплазмой. Различают два вида
эндоплазматического ретикулума: гладкий и шероховатый.
На шероховатом расположены многочисленные гранулы, представляющие собой рибосомы.
Они находятся снаружи полости, с цитоплазматической стороны, и синтезируют белки, которые
по специальному каналу сразу направляются в полость ретикулума или встраиваются в его
мембрану.
На гладком ретикулуме расположены ферменты, синтезирующие мембранные липиды.
Таким образом, эндоплазматический ретикулум образует все компоненты, нужные для
образования мембран (то есть роста их площади). От эндоплазматического ретикулума
отделяются мембранные пузырьки, внутри которых белки, синтезированные на шероховатом
ретикулуме, переносятся в следующую органеллу — аппарат, или комплекс, Гольджи.
21
22.
Комплекс ГольджиАппарат, или комплекс, Гольджи — система уплощенных мембранных
цистерн, основная функция которых — сортировка и модификация (прежде
всего гликозилирование) белков, направляемых на экспорт из клетки или
встроенных в мембрану.
Каждая группа белков, синтезированных на шероховатом ретикулуме,
собирается в определенном участке на периферии аппарата Гольджи. В
этих участках от него отделяются мембранные пузырьки, часть из которых
дает начало клеточным органеллам, таким как лизосомы. Другая
направляется к цитоплазматической мембране, сливается с ней и выделяет
свое содержимое наружу.
Таким образом осуществляется секреция из клетки таких белков, как
пищеварительные ферменты, гормоны, белки межклеточного матрикса и
гликокаликса.
22
23.
ЛизосомыЛизосомы представляют собой мембранные пузырьки, внутри которых находятся
гидролитические ферменты, расщепляющие белки, жиры, полисахариды. В
лизосомах кислая среда (рН 4,5–5,0), что отличает их от других органелл клетки.
Эта среда создается действием специального фермента — Н+- АТФазы,
перекачивающей протоны из цитоплазмы в лизосомы.
Лизосомы выполняют функцию клеточного пищеварения, расщепляя
отработавшие компоненты клетки или вещества, поглощенные в результате
фагоцитоза и пиноцитоза.
23
24.
ПероксисомыПероксисомы среди одномембранных органелл стоят особняком, т. к. способны
делиться самостоятельно, хотя не содержат ДНК. В них находятся ферменты,
катализирующие некоторые окислительно-восстановительные реакции, в которых
участвуют перекиси. Они также играют важную роль в обезвреживании многих
токсичных веществ. Белки, которые находятся в пероксисомах, поступают туда из
ЭПС и кодируются в геноме ядра. На электронных микрофотографиях пероксисом
часто можно видеть в них кристаллы ферментов.
24
25.
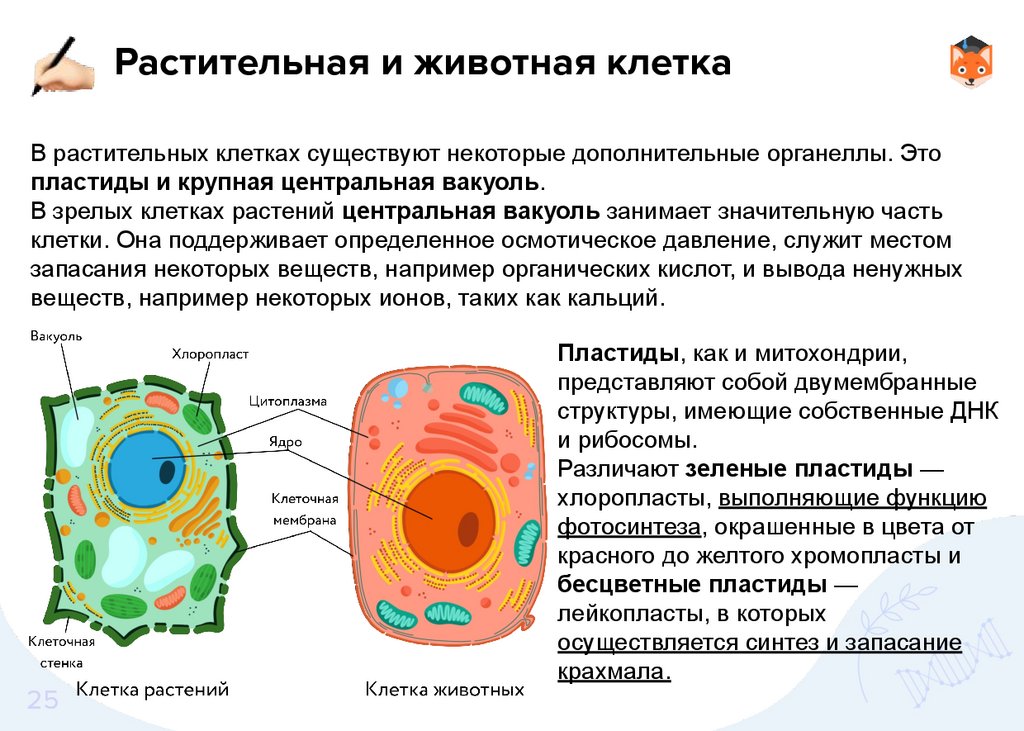
Растительная и животная клеткаВ растительных клетках существуют некоторые дополнительные органеллы. Это
пластиды и крупная центральная вакуоль.
В зрелых клетках растений центральная вакуоль занимает значительную часть
клетки. Она поддерживает определенное осмотическое давление, служит местом
запасания некоторых веществ, например органических кислот, и вывода ненужных
веществ, например некоторых ионов, таких как кальций.
Пластиды, как и митохондрии,
представляют собой двумембранные
структуры, имеющие собственные ДНК
и рибосомы.
Различают зеленые пластиды —
хлоропласты, выполняющие функцию
фотосинтеза, окрашенные в цвета от
красного до желтого хромопласты и
бесцветные пластиды —
лейкопласты, в которых
осуществляется синтез и запасание
крахмала.
25
26.
Обмен веществ. Энергетический обмен. РольАТФ
Совокупность реакций обмена веществ, протекающих в организме,
называется метаболизмом. Процессы синтеза специфических
собственных веществ из более простых называется анаболизмом, или
ассимиляцией, или пластическим обменом. В результате анаболизма
образуются ферменты, вещества, из которых построены клеточные
структуры, и т.п. Этот процесс, как правило, сопровождается большим
потреблением энергии.
26
27.
Обмен веществ. Энергетический обмен. РольАТФ
Эта энергия получается организмом в других реакциях, в которых
более сложные вещества расщепляются до простых. Эти процессы
называются катаболизмом, или диссимиляцией, или энергетическим
обменом.
Продуктами катаболизма у аэробных организмов являются СО2,
Н2О, АТФ и восстановленные переносчики водорода (НАД∙Н и
НАДФ∙Н), которые принимают атомы водорода, отщепляемые от
органических веществ в процессах окисления. Некоторые
низкомолекулярные вещества, которые образуются в ходе
катаболизма, в дальнейшем могут служить предшественниками
необходимых клетке веществ (пересечение катаболизма и анаболизма)
27
28.
Обмен веществ. Энергетический обмен. РольАТФ
Катаболизм и анаболизм тесно связаны:
анаболизм использует энергию и восстановители,
образующиеся в реакциях катаболизма, а катаболизм
осуществляется под действием ферментов, образующихся в
результате реакций анаболизма. Как правило, катаболизм
сопровождается окислением используемых веществ, а
анаболизм — восстановлением.
28
29.
Обмен веществ. Энергетический обмен. РольАТФ
29
30.
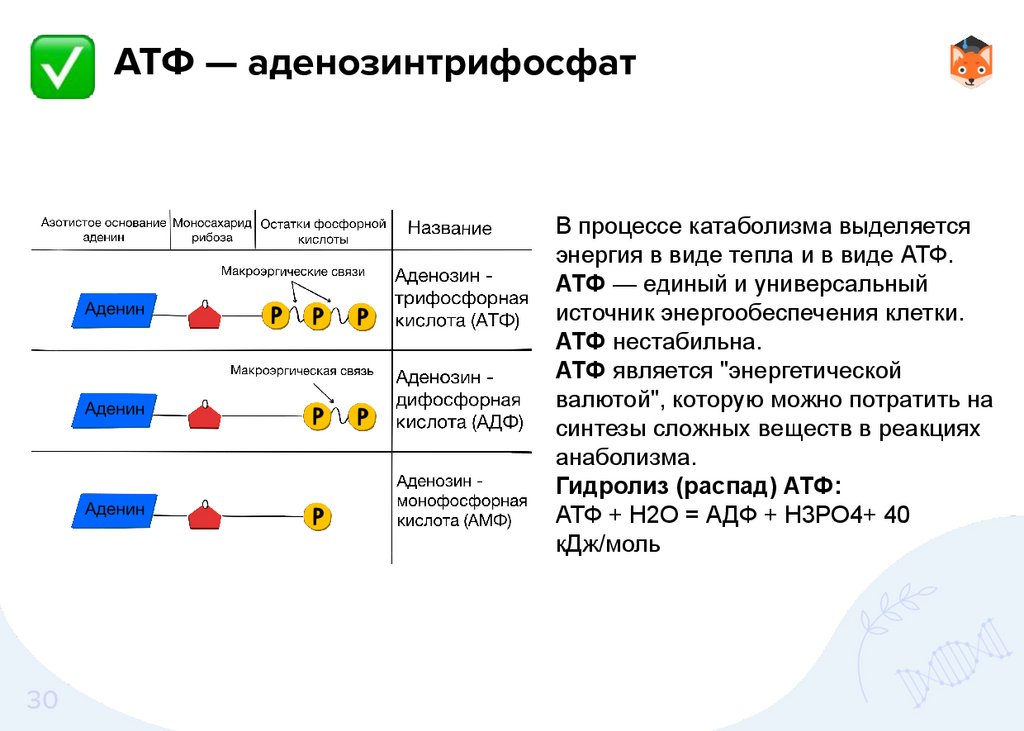
АТФ — аденозинтрифосфатВ процессе катаболизма выделяется
энергия в виде тепла и в виде АТФ.
АТФ — единый и универсальный
источник энергообеспечения клетки.
АТФ нестабильна.
АТФ является "энергетической
валютой", которую можно потратить на
синтезы сложных веществ в реакциях
анаболизма.
Гидролиз (распад) АТФ:
АТФ + Н2О = АДФ + Н3РО4+ 40
кДж/моль
30
31.
Энергетический обменЖивые организмы получают энергию в
результате окисления органических
соединений.
Окисление — процесс отдачи электронов.
Расход полученной энергии:
❏ 50% энергии выделяется в виде
тепла в окружающую среду;
❏ 50% энергии идет на пластический
обмен (синтез веществ).
В клетках растений:
крахмал → глюкоза → АТФ
В клетках животных:
гликоген → глюкоза → АТФ
31
32.
Энергетический обменФерментативное расщепление сложных органических веществ до простых в
пищеварительной системе:
➔
➔
➔
белковые молекулы — до аминокислот
липиды — до глицерина и жирных кислот
углеводы — до глюкозы
Распад (гидролиз) высокомолекулярных органических соединений
осуществляется или ферментами желудочно-кишечного тракта или
ферментами лизосом. Вся высвобождающаяся при этом энергия
рассеивается в виде тепла.
Простые вещества всасываются ворсинками тонкого кишечника:
аминокислоты и глюкоза — в кровь;
жирные кислоты и глицерин — в лимфу;
и переносятся к клеткам тканей организма.
32
33.
Энергетический обменГликолиз — расщепление глюкозы с помощью ферментов. Идет в
цитоплазме, без кислорода.
Во время этого процесса происходит дегидрирование глюкозы, акцептором
водорода служит кофермент НАД+ (никотинамидадениндинуклеотид).
Глюкоза в результате цепочки ферментативных реакций
превращается в две молекулы пировиноградной кислоты (ПВК), при
этом суммарно образуются 2 молекулы АТФ и восстановленная форма
переносчика водорода НАД·Н2.
Дальнейшая судьба ПВК зависит от присутствия кислорода в клетке: если
кислорода нет, у дрожжей и растений происходит спиртовое брожение, при
котором сначала происходит образование уксусного альдегида, а затем
этилового спирта.
У животных и некоторых бактерий при недостатке кислорода происходит
молочнокислое брожение с образованием молочной кислоты.
33
34.
Энергетический обменОкислительное фосфорилирование — процесс синтеза АТФ с участием
кислорода. Идет на мембранах крист митохондрий в присутствии
кислорода.
34
Пировиноградная кислота, образовавшаяся при бескислородном
расщеплении глюкозы, окисляется до конечных продуктов СО2 и Н2О. Этот
многоступенчатый ферментативный процесс называется циклом Кребса, или
циклом трикарбоновых кислот.
В результате клеточного дыхания при распаде двух молекул пировиноградной
кислоты синтезируются 36 молекул АТФ:
2С3Н4О3 + 32О2 + 36АДФ + 36Н3РО4 → 6СО2 + 58Н2О + 36АТФ.
Кроме того, нужно помнить, что две молекулы АТФ запасаются в ходе
бескислородного расщепления каждой молекулы глюкозы. Суммарная реакция
расщепления глюкозы до углекислого газа и воды выглядит следующим
образом:
С6Н12О6 + 6О2 + 38АДФ → 6СО2 + 6Н2О + 38АТФ + Qт, где Qт — тепловая
энергия.
Таким образом при окислительном фосфорилировании образуется в 18 раз
больше энергии (36 АТФ), чем при гликолизе (2 АТФ). Гликолиз используют
некоторые бактерии и паразиты, обитающие в анаэробных условиях.
35.
Окислительное декарбоксилированиепирувата
На первом этапе в матриксе митохондрий от пировиноградной кислоты (С3)
отщепляется молекула CO2, при этом образуется остаток уксусной кислоты
(ацетил), который присоединяется к специальной молекуле-переносчику —
коферменту А (КоА). Образуется ацетил-КоА. Он несет два атома С в виде
ацетильного остатка, так как один из атомов С3-соединения — пирувата — уже
ушёл в виде CO2. В ацетил-КоА превращаются при окислении в митохондриях и
жирные кислоты, и ряд аминокислот. Это окислительный процесс, при этом
образуется 1 молекула восстановленного НАДН.
35
36.
Цикл КребсаДальнейший путь окисления является общим практически для всех
энергетических субстратов, а не только для углеводов. Он носит
название цикл ди- и трикарбоновых кислот или, по имени его
открывателя, цикл Кребса.
Первоначально двухуглеродный (С2) ацетильный остаток из
ацетил-КоА переносится на щавелевоуксусную кислоту (С4) с
образованием 6-углеродной (С6) лимонной кислоты. Она
окисляется в ряде последовательных реакций путём переноса её
водородов на молекулы НАД+ или других переносчиков и
отщепления окисленных атомов углерода в виде 2 молекул СО2 .
В ходе этого процесса синтезируется ещё 1 молекула АТФ или
ГТФ. В конце этого процесса образуется молекула
щавелевоуксусной кислоты, которая снова вступает в цикл.
36
37.
Цикл КребсаТаким образом, продуктами цикла Кребса являются:
❏ 2 молекулы углекислого газа
❏ 3 молекулы НАД∙Н
❏ 1 молекула восстановленного переносчика, называемого ФАД∙Н 2
Кроме того, по одной молекуле НАД∙Н образовалось при окислении в
гликолизе и при окислении пировиноградной кислоты в ацетил-КоА. Так как
при окислении 1 молекулы глюкозы образовалось две молекулы пирувата,
это количество надо умножить на два.
Таким образом, полное аэробное окисление 1 молекулы глюкозы
приводит к образованию:
❏ 10 молекул НАД∙Н
❏ 2 молекул ФАД∙Н2
❏ 4 молекул АТФ
Результатом цикла Кребса является полное окисление углеродного скелета
пищевого субстрата до CO2 и H2O. Катаболизм органических соединений
на этом завершён. Однако остается задача окисления переносчиков
водорода (НАДН), накопивших водород, и получения энергии в виде АТФ.
37
38.
Цикл Кребса38
39.
Транспорт электронов и синтез АТФНа внутренней мембране митохондрии атомы водорода, накопившиеся в
процессе биологического окисления на молекулах переносчиков, отдают свои
электроны, превращаясь в протоны. Электроны через цепь специальных
белков переносятся на молекулярный кислород, в результате чего образуется
вода. Система белков-переносчиков электронов называется
электронтранспортной цепью, а сами такие белки — цитохромами.
Присоединяя электроны из электронтранспортной цепи и ионы водорода из
среды, молекулярный кислород превращается в воду.
39
40.
Транспорт электронов и синтез АТФПеренос электронов по электронтранспортной цепи
сопровождается переносом ионов водорода через
внутреннюю мембрану митохондрии из матрикса в
межмембранное пространство. Окисление одной
молекулы НАД∙Н приводит к переносу 10 ионов водорода.
В результате на внутренней мембране митохондрий
формируется разность потенциалов (протоны
скопились в межмембранном пространстве).
Таким образом, энергия восстановленных переносчиков
превращается в электрическую энергию — энергию
мембранного потенциала. Это важнейшая форма
существования энергии в клетке, наряду с энергией
химических связей и тепловой энергией.
40
41.
Транспорт электронов и синтез АТФСинтез АТФ
Превращение энергии
мембранного потенциала в
энергию АТФ осуществляет
сложный белковый комплекс,
называемый АТФ-синтазой. На
электронных микрофотографиях
высокого разрешения АТФсинтаза видна на кристах
митохондрий в виде грибовидных
тел.
41
42.
Транспорт электронов и синтез АТФСинтез АТФ
В этом комплексе есть канал, по которому ионы водорода по электрическому
потенциалу проходят внутрь (в матрикс) митохондрии из межмембранного
пространства, а энергия их потока используется для обеспечения реакции
присоединения фосфата к АДФ с образованием АТФ (фосфорилирование
АТФ). Это происходит подобно тому, как на гидроэлектростанциях энергия
пассивно падающей с высоты плотины воды, которая движет лопасти турбин,
превращается в электрическую энергию.
Для синтеза одной молекулы АТФ необходимо перенести три протона,
поэтому окисление 1 молекулы НАД∙Н даёт три молекулы АТФ, одной
молекулы ФАД∙Н2 – 2 молекулы АТФ. Суммарно окисление одной молекулы
глюкозы может дать 38 молекул АТФ. Часть высвобожденной энергии
выделяется в виде тепла.
42