


















































 biology
biologySimilar presentations:





")




Мерцательный эпителий. Регуляция дыхания
1.
Мерцательный эпителийПрофессор Берг М.Д.
2. Регуляция дыхания
Берг М.Д.Регуляция дыхания
3.
План лекции1. Клеточный уровень управления дыханием. Роль HIF-1a.
2. Дыхательный центр, его отделы и функции. Методы исследования.
3. Жизненно важный отдел. Структура и функции центра вдоха,
центра выдоха, пневмотаксического центра.
4. Современные представления о механизме периодичности дыхания.
Генератор центрального инспираторного возбуждения,
механизм обрывания вдоха, механизм плавной смены вдоха и выдоха.
5. Локализация и функции спинального отдела дыхательного центра.
6. Локализация и функции адаптирующих центров.
7. Хеморецепторы и их роль в регуляции дыхания. Периферические
и центральные хеморецепторы.
8. Рефлекторные влияния на дыхание с рецепторов легких,
воздухоносных путей и дыхательных мышц.
9. Функциональная система дыхания.
10. Дыхание плода. Дыхание при разных физиологических состояниях.
Профессор Берг М.Д.
4.
Клеточный уровень управления дыханиемВсе функции клетки связаны с потреблением
энергии: для сокращения, секреции,
генерации биопотенциалов, синтеза веществ,
пролиферации и т.д. Источником энергии
служит АТФ, который в больших количествах
образуется в митохондриях в процессе
окислительного фосфорилирования глюкозы
и жирных кислот. Количество кислорода,
идущего на эти цели, должно соответствовать
потребностям клетки.
Профессор Берг М.Д.
5.
За исследование ферментов клеточногодыхания немецкий биохимик Отто Варбург
получил Нобелевскую премию в 1931 году.
Однако, как клетка отслеживает
концентрацию кислорода при меняющейся
потребности в нем, стало известно не так
давно. В 2019 году была присуждена
Нобелевская премия за исследование
клеточных механизмов чувствительности
к гипоксии. Её удостоились Грегг Семенза
(Gregg L. Semensa), Питер Рэтклифф (Peter
J.Ratcliff ), Уильям Келин-младший (Willian
Kaelin, Jr.).
Профессор Берг М.Д.
6.
Работами не только этих ученых и ихсотрудников, но и многих других было
установлено, что механизм
чувствительности к гипоксии работает с
более чем 300 генов. От этого зависит и
работа мышц, и рост кровеносных
сосудов, и работа нейронов, иммунные
реакции, развитие эмбриона и др.
Сигнальной молекулой служит HIF-1а
(hypokia-inducible factor, гипоксией
индуцированный фактор), который
чувствует уровень кислорода в клетке.
Профессор Берг М.Д.
7.
Когда кислорода мало (гипоксия), HIF-1астановится много из-за того, что перестает
работать механизм его утилизации путем
разрушения в протеасомах. При нормоксии к
HIF-1а присоединяется убиквитин – метка,
которая показывает протеасоме, что белок
не нужен и его следует утилизировать. При
нормальном содержании кислорода на
белке HIF-1а происходит гидроксилирование
пролина – на остатках аминокислоты
пролина в белковой молекуле появляются
гидроксильные группы ОН.
Профессор Берг М.Д.
8.
В таком модифицированном виде онвзаимодействует с белком VHL (von HippelLinau), который помогает навесить на него
убиквитин и отправить на утилизацию. Если
гипоксия, то HIF-1а остается невидимым для
протеасомы из-за отсутствия метки VHL.
HIF состоит из двух частей: уровень
кислорода чувствует HIF-1а, а второй белок
– ARNT – нужен, чтобы посадить HIF-1а на
ДНК и простимулировать тем самым ген,
отвечающий за ту или иную функцию.
Профессор Берг М.Д.
9.
Как ген может почувствовать недостатоккислорода? Генетическую информацию с
ДНК считывают транскрипционные
ферменты, их активность можно усилить или
ослабить. Это делают белки «факторы
транскрипции». Рядом с геном всегда есть
специальные регуляторные участки ДНК,
которые служат посадочными площадками
для факторов транскрипции: сев на ДНК, они
либо привлекают к гену больше
транскрипционных ферментов (ген
становится активнее), либо не дают им
работать с этим геном. Таким фактором
транскрипции является HIF-1а.
Профессор Берг М.Д.
10.
Для управления внутриклеточнымипроцессами, которые зависят от
потребления кислорода, организм должен
поддерживать нормоксию тканевой
жидкости и крови, а значит управлять
поступлением кислорода из внешней
среды. Это достигается ритмическим
чередованием вдоха и выдоха и
изменением глубины и частоты дыхания.
Поэтому на уровне организма регуляция
дыхания связана с функцией
дыхательного центра.
Профессор Берг М.Д.
11.
Дыхательный центр.Отделы и их функции.
Регуляция дыхания – управление
ритмичным чередованием вдоха и
выдоха и их глубиной с целью поддержания
постоянства газового состава крови при
различных состояниях организма.
Эту функцию выполняет дыхательный
центр.
Профессор Берг М.Д.
12.
Его отделы:1. Жизненно важный отдел (собственно
дыхательный центр) - бульбопонтийный
отдел - БПО. Смена вдоха и выдоха.
Плавный переход с одной фазы дыхания
на другую. Изменение глубины и частоты
дыхания.
2. Исполнительный отдел (спинальный
отдел ДЦ). Исполнить команды БПО и ОП.
3. Отдел приспособления к меняющимся
условиям (ЛС, гипоталамус, КБП) - ОП.
Приспособить транспорт О2 и энергообмен
к потребностям организма.
Профессор Берг М.Д.
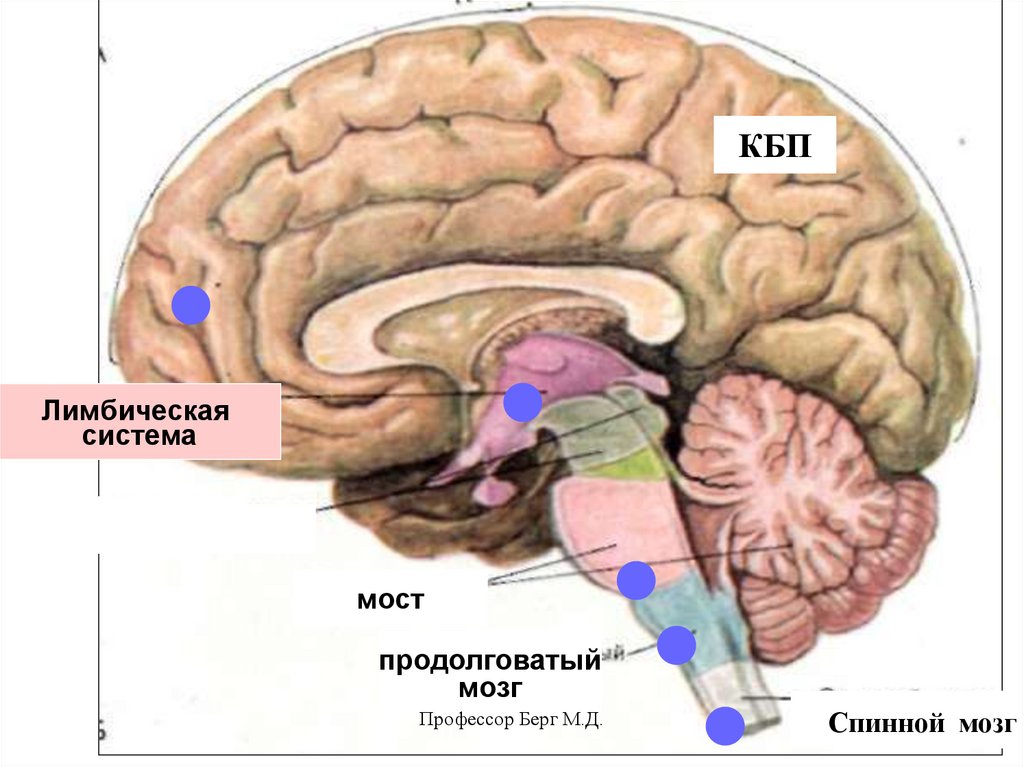
13.
КБПЛимбическая
система
мост
продолговатый
мозг
Профессор Берг М.Д.
Спинной мозг
14.
Методы исследования дыхательногоцентра у животных:
1. Перерезки мозга на разных уровнях.
2. Раздражение структур мозга.
Жизненно важный бульбо-понтийный
отдел включает:
• дорзальные и вентральные ядра
продолговатого мозга,
• медиальное парабрахиальное ядро
моста.
Профессор Берг М.Д.
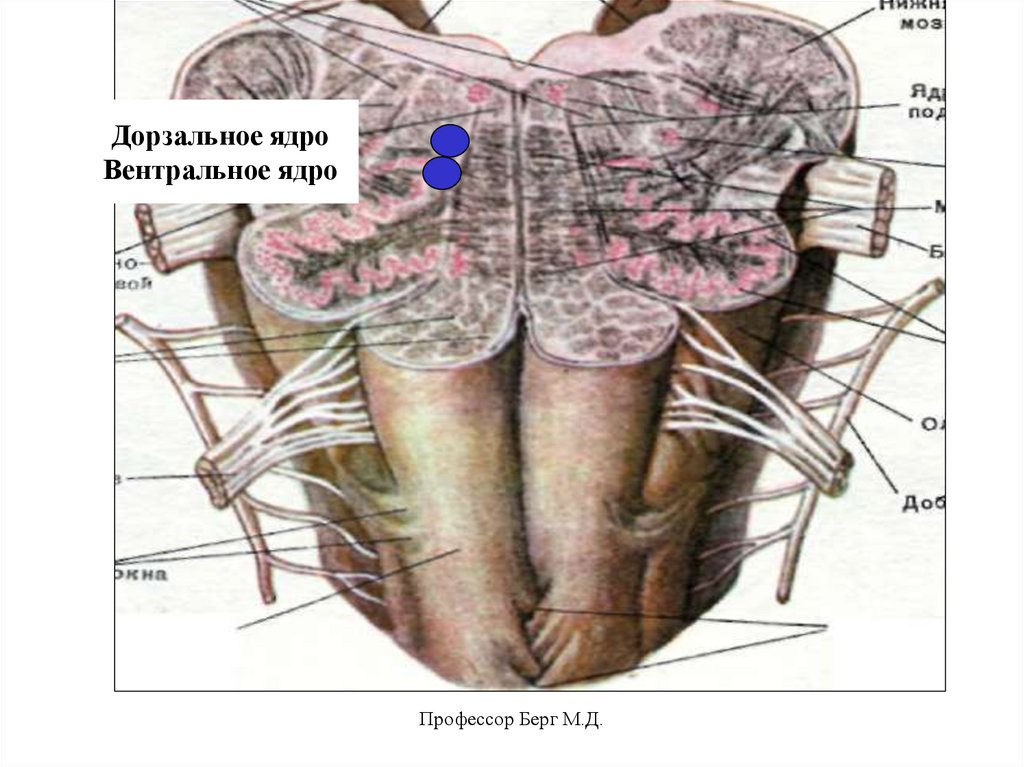
15.
Дорзальное ядроВентральное ядро
Профессор Берг М.Д.
16.
Дорзальное ядро:• управляет сокращением диафрагмы
через альфа-мотонейроны С 3-4 (инспирация
– вдох).
Вентральное ядро:
• управляет сокращением наружных межреберных мышц через альфа- мотонейроны
Т1-6 (инспирация),
• управляет сокращением внутренних межреберных мышц через альфа- мотонейроны
Т1-6 (экспирация – выдох),
• управляет сокращением мышц живота
через альфа-мотонейроны Т9-12, L1-5
(форсированная экспирация).
Профессор Берг М.Д.
17.
Нейроны этих ядер объединяютфункционально в:
1) центр вдоха
(инспираторный центр), в него входят
нейроны, управляющие мышцами вдоха.
2) центр выдоха
(экспираторный центр), в него входят
нейроны, тормозящие центр вдоха,
и нейроны, управляющие мышцами
выдоха.
Профессор Берг М.Д.
18.
Функции центра вдоха:• восприятие сигналов от хеморецепторов,
• генерация вдоха,
• передача сигнала к мотонейронам мышц
вдоха,
• возбуждение центра выдоха через
нейроны пневмотаксического центра
моста.
Профессор Берг М.Д.
19.
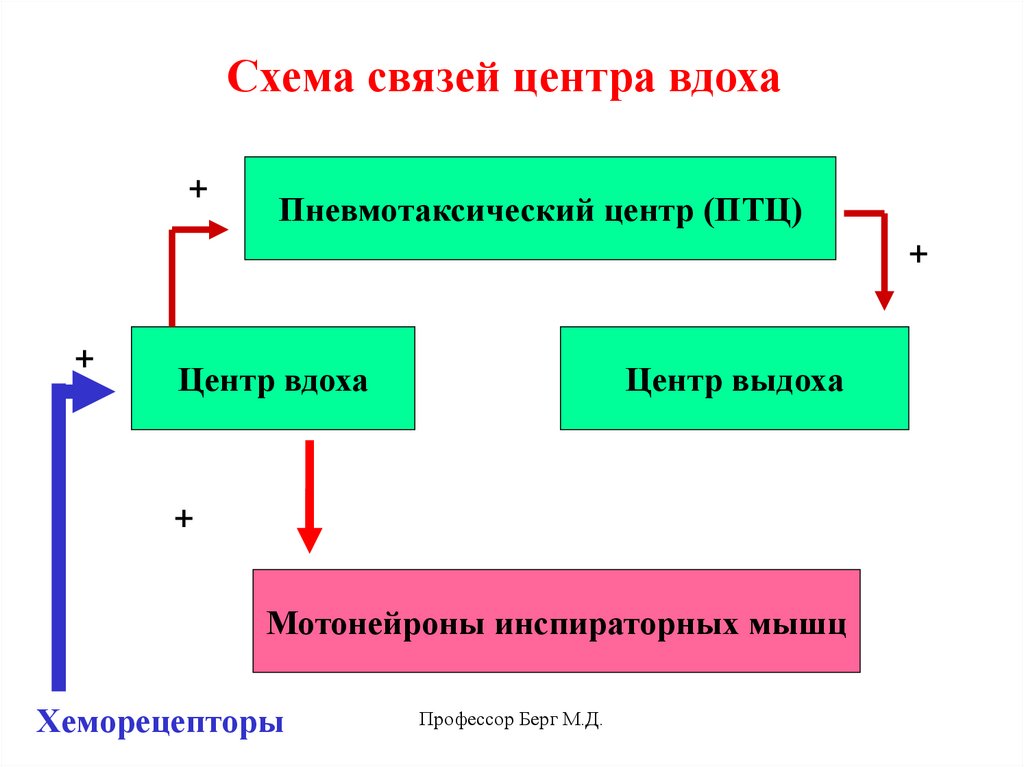
Схема связей центра вдоха+
Пневмотаксический центр (ПТЦ)
+
+
Центр вдоха
Центр выдоха
+
Мотонейроны инспираторных мышц
Хеморецепторы
Профессор Берг М.Д.
20.
Функции центра выдоха:• восприятие сигналов от механорецепторов
легких,
• торможение инспираторного отдела
(центра вдоха),
• передача сигнала на мотонейроны мышц
выдоха для форсированного дыхания.
Профессор Берг М.Д.
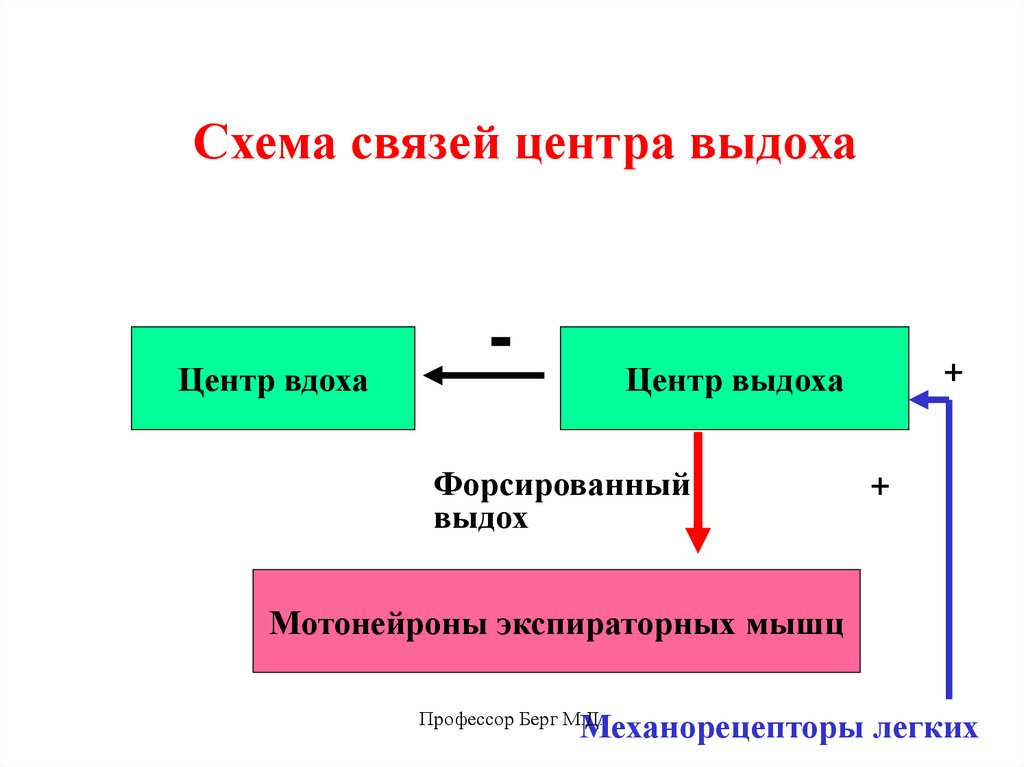
21.
Схема связей центра выдохаЦентр вдоха
-
+
Центр выдоха
Форсированный
выдох
+
Мотонейроны экспираторных мышц
Механорецепторы легких
Профессор Берг М.Д.
22.
Медиальное парабрахиальное ядро мостаназывают пневмотаксическим центром ПТЦ.
Его функции:
• плавная смена вдоха и выдоха,
• повышение возбудимости механизма
выключения вдоха.
Он получает информацию от центра
вдоха и передает ее центру выдоха.
Профессор Берг М.Д.
23.
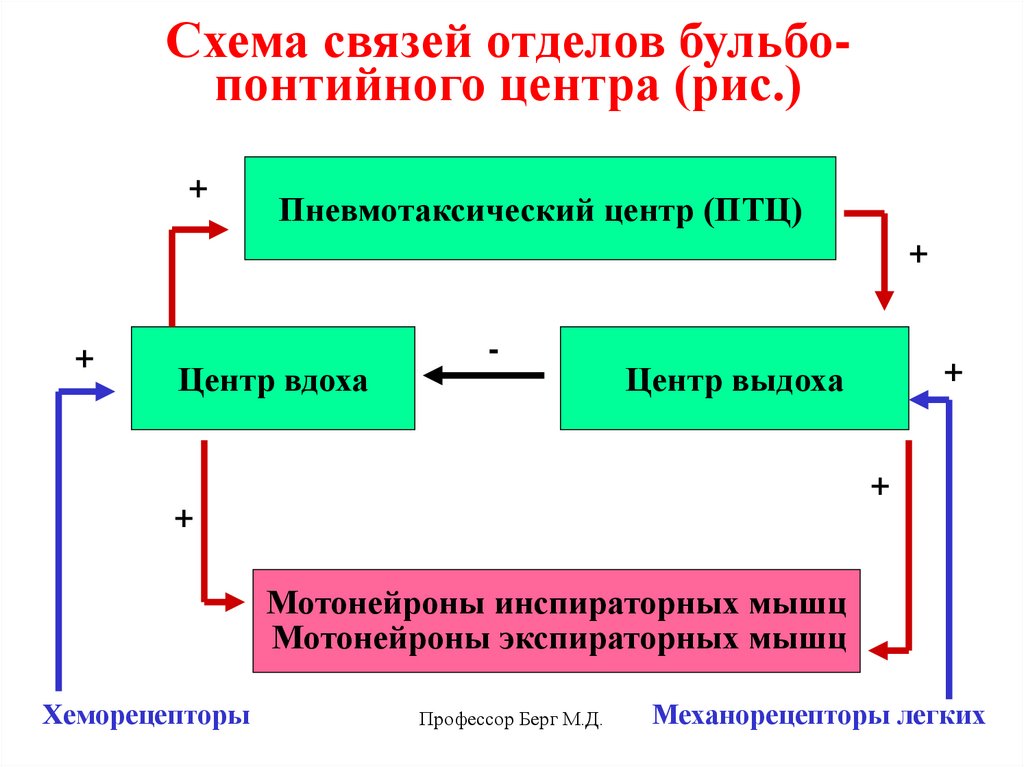
Схема связей отделов бульбопонтийного центра (рис.)+
Пневмотаксический центр (ПТЦ)
+
+
Центр вдоха
-
+
Центр выдоха
+
+
Мотонейроны инспираторных мышц
Мотонейроны экспираторных мышц
Хеморецепторы
Профессор Берг М.Д.
Механорецепторы легких
24.
Современные представления омеханизме периодичности
дыхания
Профессор Берг М.Д.
25.
В ДЦ выделяют 3 механизма, которыеуправляют нормальным дыханием:
1. Генератор центрального инспираторного возбуждения.
2. Механизм выключения инспирации.
3. Механизм плавной смены вдоха и
выдоха.
Профессор Берг М.Д.
26.
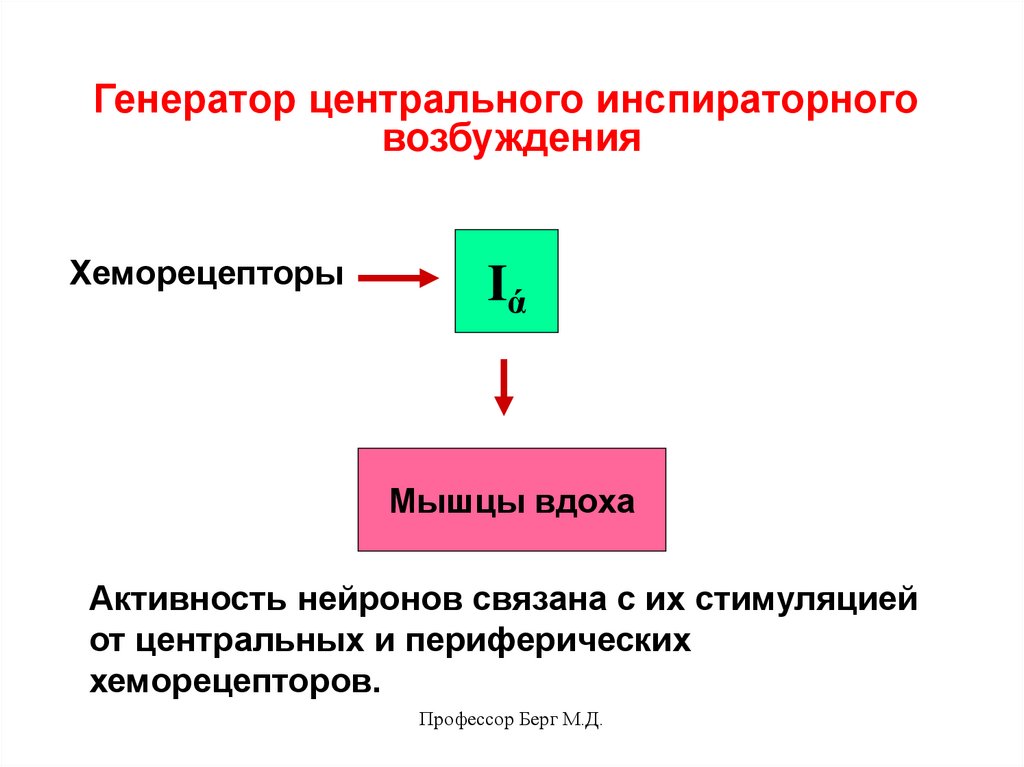
Генератор центрального инспираторноговозбуждения
К нему относят инспираторные нейроны
(ранние и полные) - нейроны Iα . Они
проявляют постоянную активность,
т.е. генерируют ПД, которые возбуждают
нейроны дорзального и вентрального
ядер, управляющие диафрагмой и
наружными межреберными мышцами,
т.е. мышцами вдоха.
Профессор Берг М.Д.
27.
Генератор центрального инспираторноговозбуждения
Хеморецепторы
Iά
Мышцы вдоха
Активность нейронов связана с их стимуляцией
от центральных и периферических
хеморецепторов.
Профессор Берг М.Д.
28.
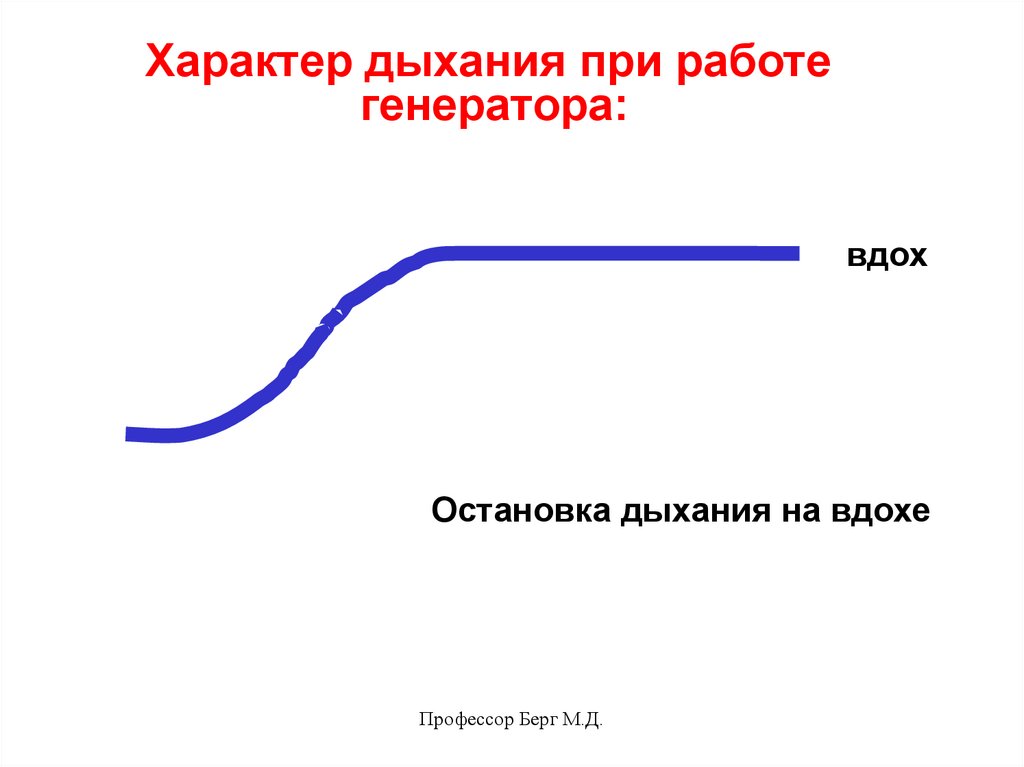
Характер дыхания при работегенератора:
вдох
Остановка дыхания на вдохе
Профессор Берг М.Д.
29.
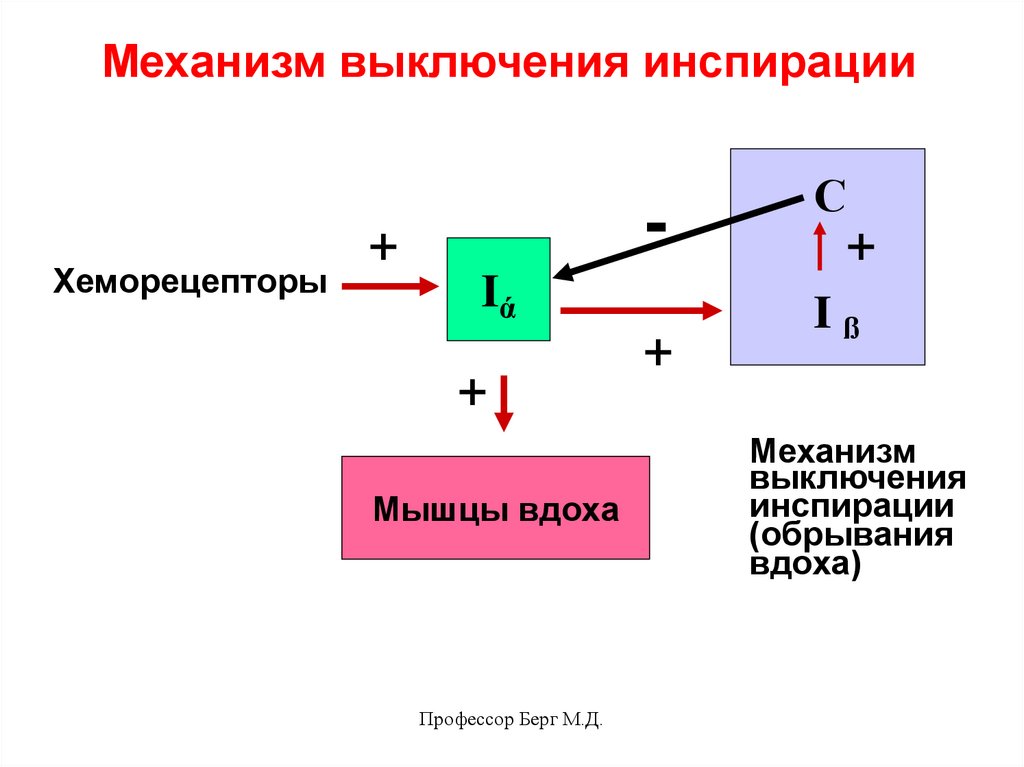
Механизм выключения инспирациипредставлен нейронами дыхательного
центра продолговатого мозга – Iß и С.
Нейроны Iß (инспираторные поздние)
возбуждаются от генератора и возбуждают
нейроны С, которые являются тормозными. Они быстро тормозят активность
генератора, наступает выдох.
Их связи с генератором центрального
инспираторного возбуждения
представлены на схеме:
Профессор Берг М.Д.
30.
Механизм выключения инспирацииХеморецепторы
-
+
Iά
+
Мышцы вдоха
Профессор Берг М.Д.
+
С
+
Iß
Механизм
выключения
инспирации
(обрывания
вдоха)
31.
Тормозные связи между нейронамиработают через глицин и ГАМК.
Глицин реализует функцию быстрого
выключения фазы дыхательного цикла –
вдоха и выдоха,
ГАМК – медленного, до окончания
соответствующей фазы дыхания.
ГАМК-рецепторы двух типов: ГАМКА
открывают каналы для хлора, ГАМКВ – для
калия.
Профессор Берг М.Д.
32.
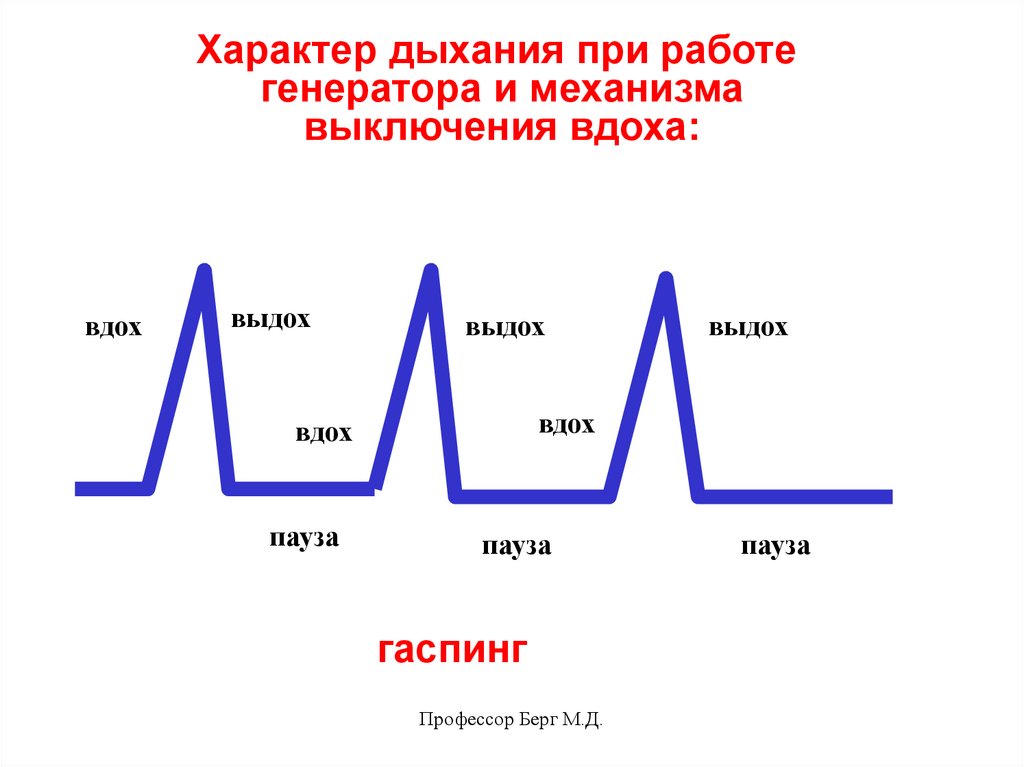
Характер дыхания при работегенератора и механизма
выключения вдоха:
вдох
выдох
выдох
вдох
вдох
пауза
выдох
пауза
гаспинг
Профессор Берг М.Д.
пауза
33.
Механизм плавной смены вдоха ивыдоха представлен 2-мя механизмами:
• нейронами ПТЦ и
• рефлексом Геринга-Брейера.
Нейроны ПТЦ возбуждаются от генератора
центрального инспираторного возбуждения
и возбуждают механизм обрывания вдоха.
Тормозные нейроны работают с медиатором
ГАМК.
Профессор Берг М.Д.
34.
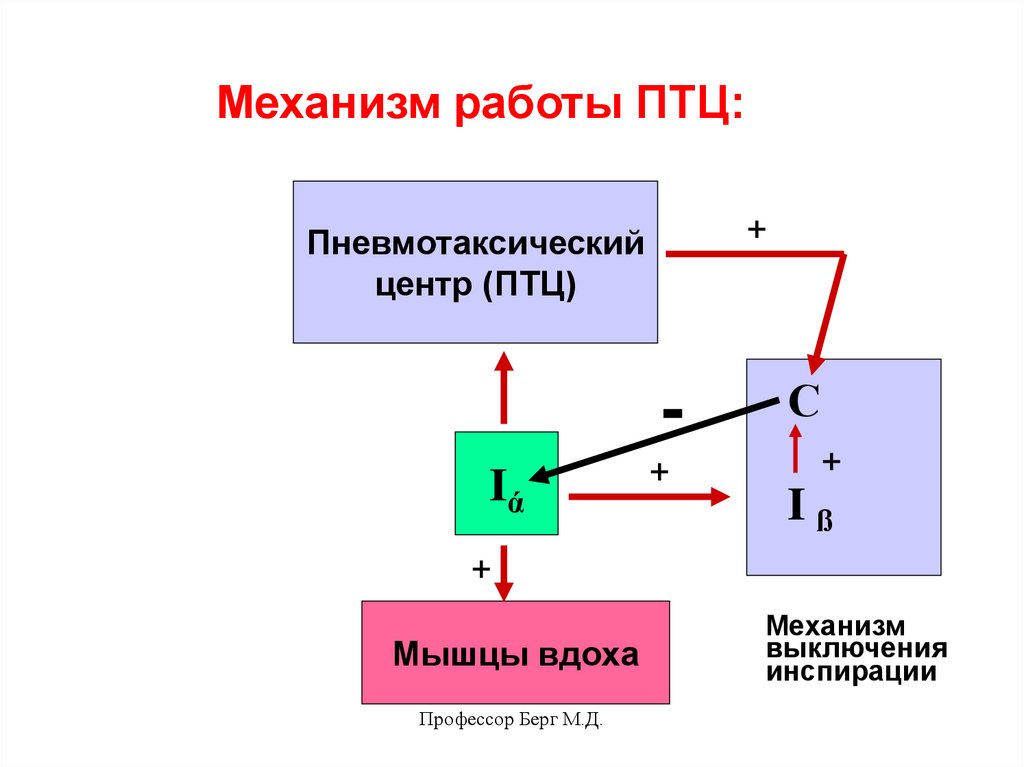
Механизм работы ПТЦ:+
Пневмотаксический
центр (ПТЦ)
Iά
+
С
+
Iß
+
Мышцы вдоха
Профессор Берг М.Д.
Механизм
выключения
инспирации
35.
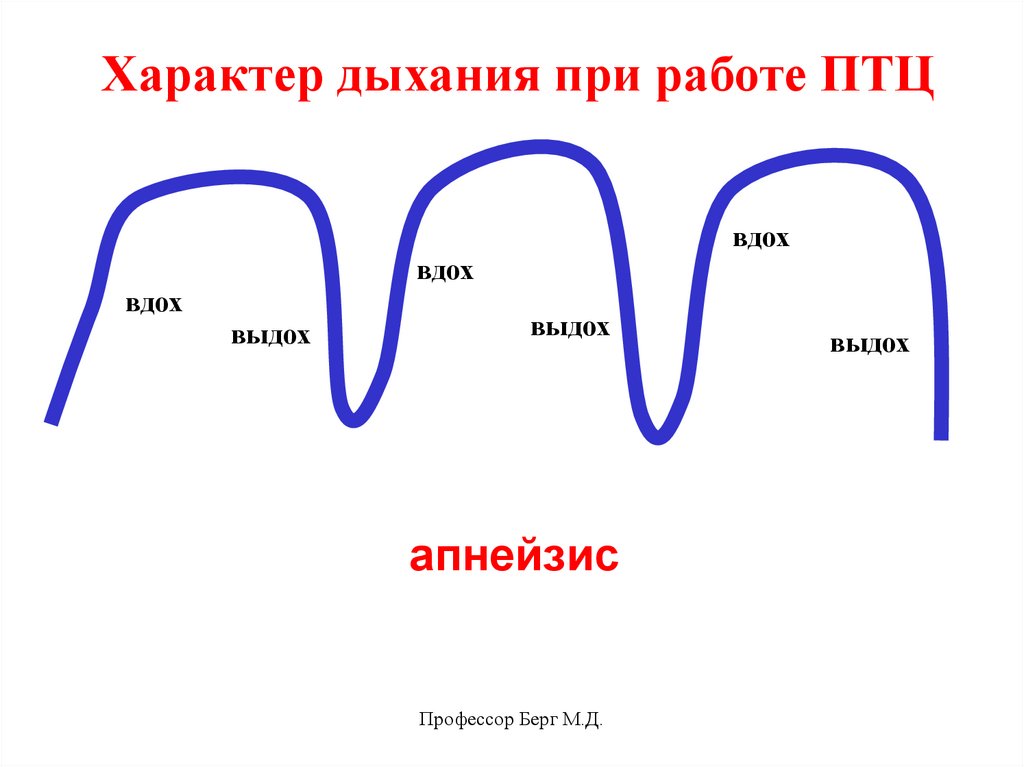
Характер дыхания при работе ПТЦвдох
вдох
вдох
выдох
выдох
апнейзис
Профессор Берг М.Д.
выдох
36.
Механизм Геринга-БрейераВ респираторном отделе легких находятся
механорецепторы – окончания дендритов
чувствительных нейронов Х пары ч/н.
Они реагируют на растяжение легких
при вдохе. Сигналы от них поступают
к нейронам Iβ . Включается механизм
обрывания вдоха. Тормозные нейроны
работают с медиатором ГАМК.
Профессор Берг М.Д.
37.
Механизм Геринга-БрейераС
Хеморецепторы
+
Iά
+
+
Iß
+
Мышцы вдоха
Механизм
выключения
инспирации
Х
+
Механорецепторы легких
(МРЛ)
Профессор Берг М.Д.
38.
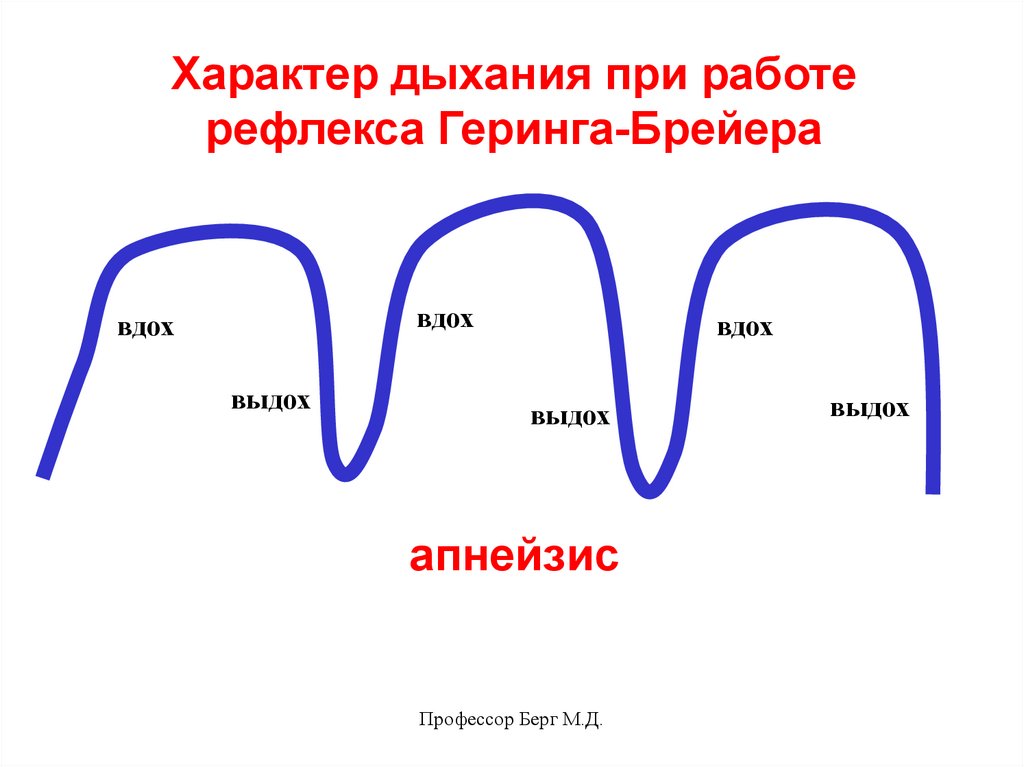
Характер дыхания при работерефлекса Геринга-Брейера
вдох
вдох
выдох
вдох
выдох
апнейзис
Профессор Берг М.Д.
выдох
39.
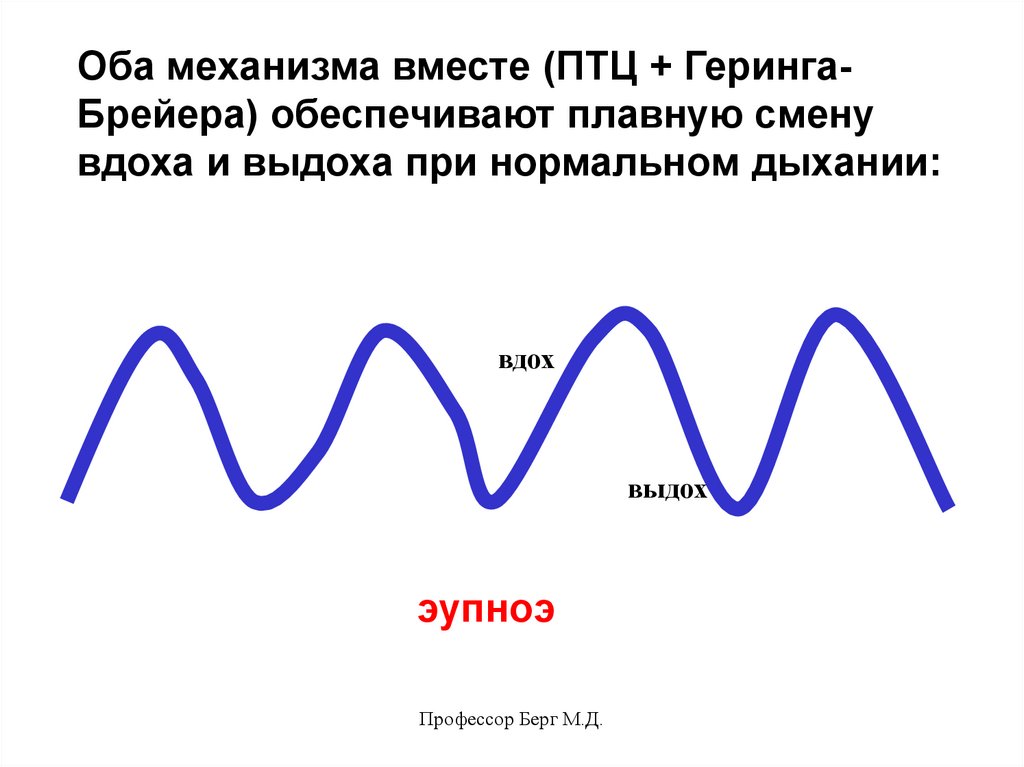
Оба механизма вместе (ПТЦ + ГерингаБрейера) обеспечивают плавную сменувдоха и выдоха при нормальном дыхании:
вдох
выдох
эупноэ
Профессор Берг М.Д.
40.
Функции спинального отдела ДЦВ передних рогах спинного мозга лежат
мотонейроны дыхательных мышц.
Сегменты С3-4 иннервируют диафрагму, Тh1-6 –
наружные межреберные мышцы.
Их функции:
• выполнение команд ДЦ,
• поддержание заданного объема дыхания
через петлю обратной связи, т.е. от
проприорецепторов дыхательных мышц.
Профессор Берг М.Д.
41.
Функции спинального отдела ДЦДЦ
Для поддержания заданной
активности мотонейронов
работает обратная связь
«-»
«+»
Профессор Берг М.Д.
Рецепторы Гольджи
Рецепторы растяжения
42.
Функции адаптирующих центровЛимбическая система и гипоталамус:
• включение ДЦ в гомеостатические
реакции (поддержание постоянства температуры крови, питательных веществ и др.),
• включение ДЦ в обеспечение врожденных
поведенческих реакций достижения цели,
• включение ДЦ в обеспечение эмоций,
• включение ДЦ в приобретенные программы
гомеостатического поведения и реакций
достижения цели.
Профессор Берг М.Д.
43.
Кора больших полушарий:• опережающая регуляция дыхания
(условный рефлекс),
• произвольная регуляция дыхания (апноэ,
гиперпноэ),
• приспособление дыхания к речи,
• субъективная оценка изменений в
дыхательной системе (трудно дышать,
задыхаюсь, тяжесть в груди и др.).
Влияние адаптирующих центров на дыхание
можно наблюдать на пневмограмме (запись
движений грудной клетки) – используется как
компонент детектора лжи.
Профессор Берг М.Д.
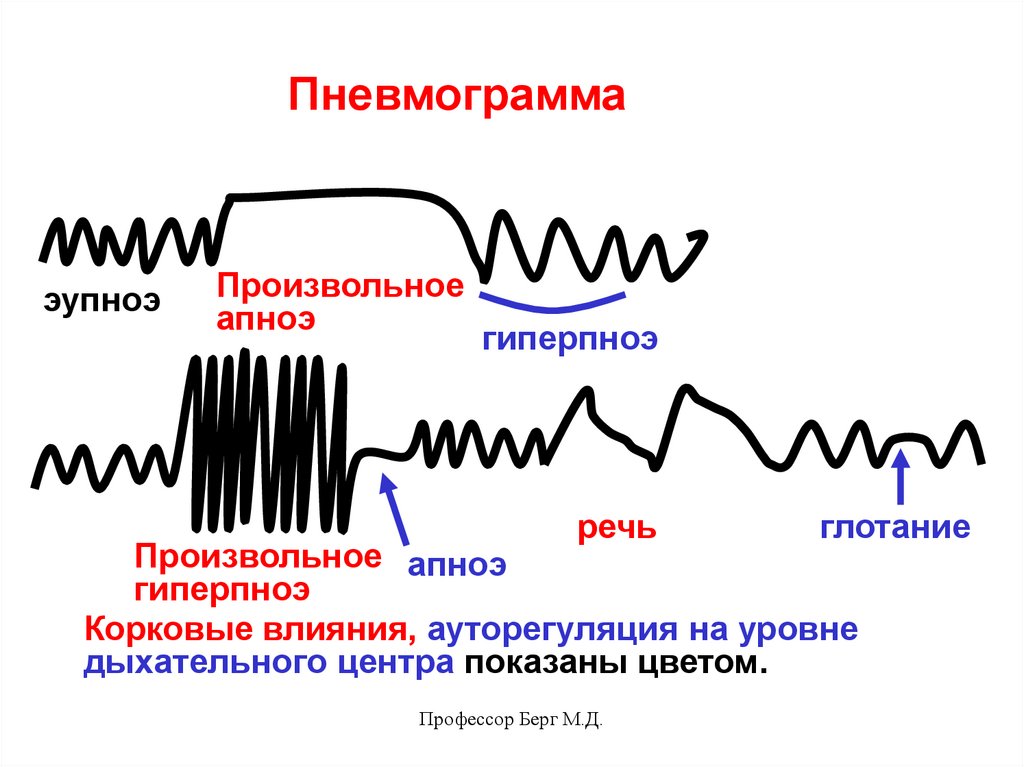
44.
Пневмограммаэупноэ
Произвольное
апноэ
гиперпноэ
речь
глотание
Произвольное апноэ
гиперпноэ
Корковые влияния, ауторегуляция на уровне
дыхательного центра показаны цветом.
Профессор Берг М.Д.
45.
Хеморецепторы и их рольв регуляции дыхания
В 1938 году за исследование механизма
работы хеморецепторов каротидного
синуса, которые реагируют на гипоксию,
дали Нобелевскую премию бельгийцу
Корнею Хеймансу.
Профессор Берг М.Д.
46.
Хеморецепторы – рецепторы, реагирующиена изменение химического состава
внутренней среды. В данном случае –
на изменение рН, рО2, рСО2.
Они регулируют:
• уровень вентиляции легких (ЧД, ДО, МОД),
• а также – МОК и
• выход эритроцитов из селезенки
(транспорт газов).
Профессор Берг М.Д.
47.
Классификация хеморецепторов:1. Периферические:
• сосудистые,
• тканевые.
2. Центральные.
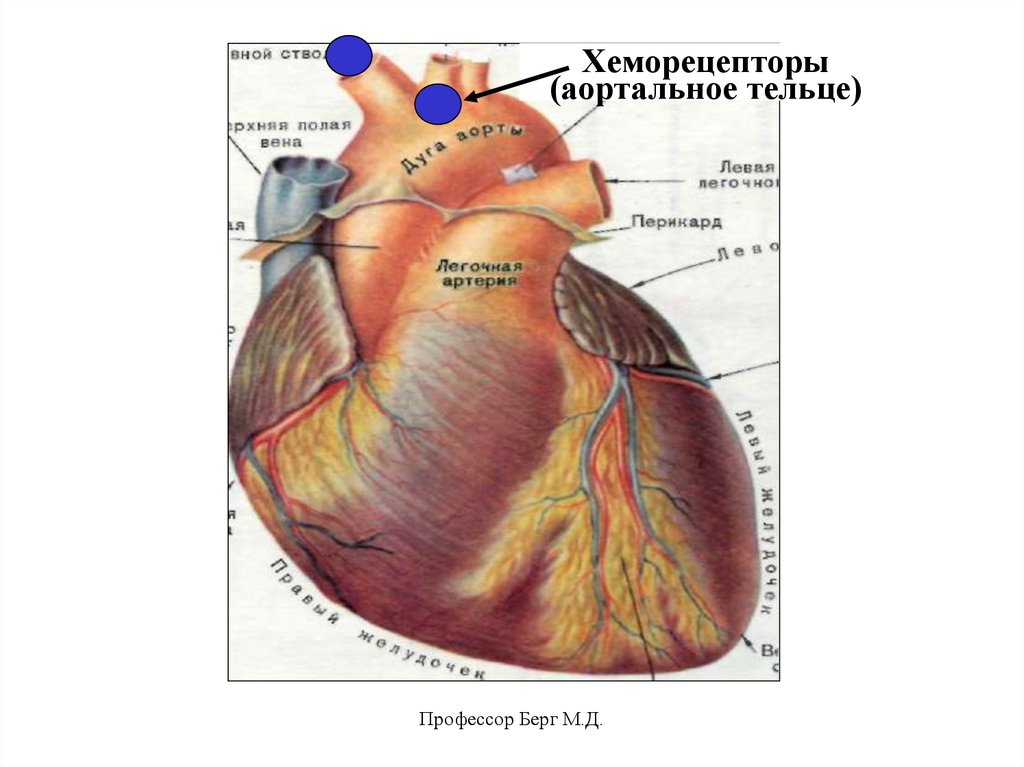
Сосудистые хеморецепторы:
• аортальные тельца (в дуге аорты) –
Х пара ч/м н.
• каротидные тельца (в каротидном
синусе) – IХ пара ч/м н.
Это вторичночувствующие рецепторы.
Профессор Берг М.Д.
48.
Хеморецепторы(аортальное тельце)
Профессор Берг М.Д.
49.
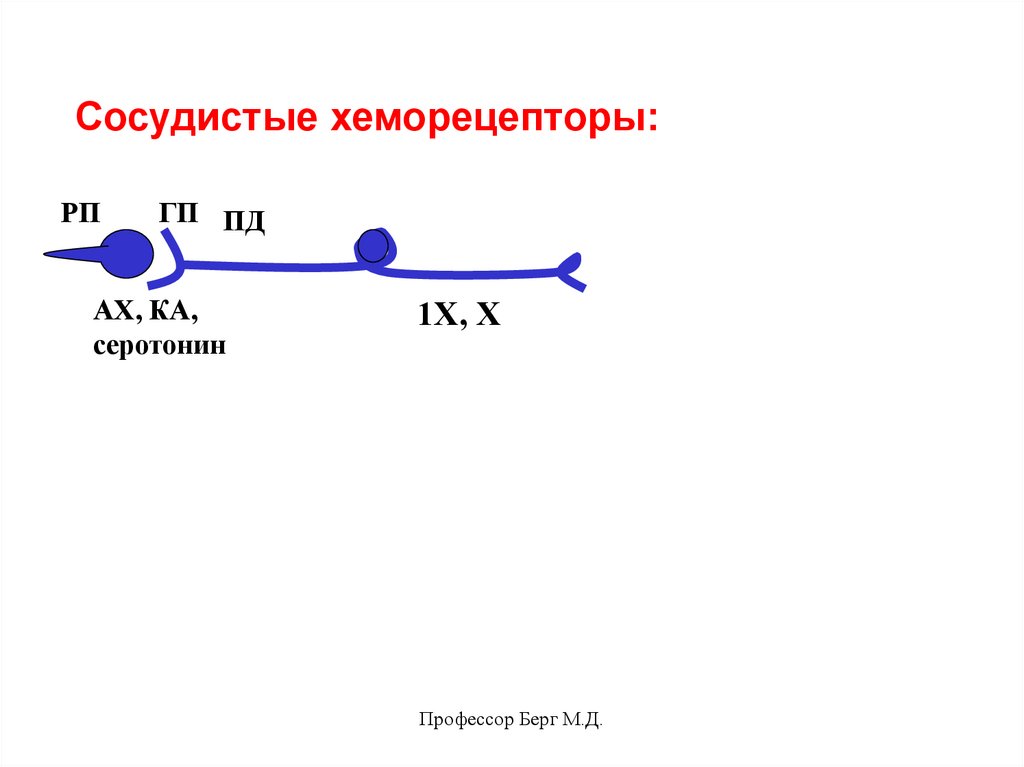
Сосудистые хеморецепторы:РП
ГП ПД
АХ, КА,
серотонин
1Х, Х
Профессор Берг М.Д.
50.
Раздражители:• снижение рО2 в артериальной крови
ниже 100 мм рт.ст. (гипоксемия),
• увеличение рСО2 в артериальной крови
выше 40 мм рт.ст. (гиперкапния),
• увеличение концентрации Н+ в артериальной крови (ацидоз).
Профессор Берг М.Д.
51.
Диапазон чувствительности к сдвигам рО2 –100-40 мм рт.ст.
Диапазон чувствительности к сдвигам рСО2 –
40-60 мм рт.ст.
Ацидоз – рН менее 7,4.
У детей до года чувствительность сосудистых
хеморецепторов снижена, что создает опасность
внезапной смерти в ночное время
(«присыпание»).
Профессор Берг М.Д.
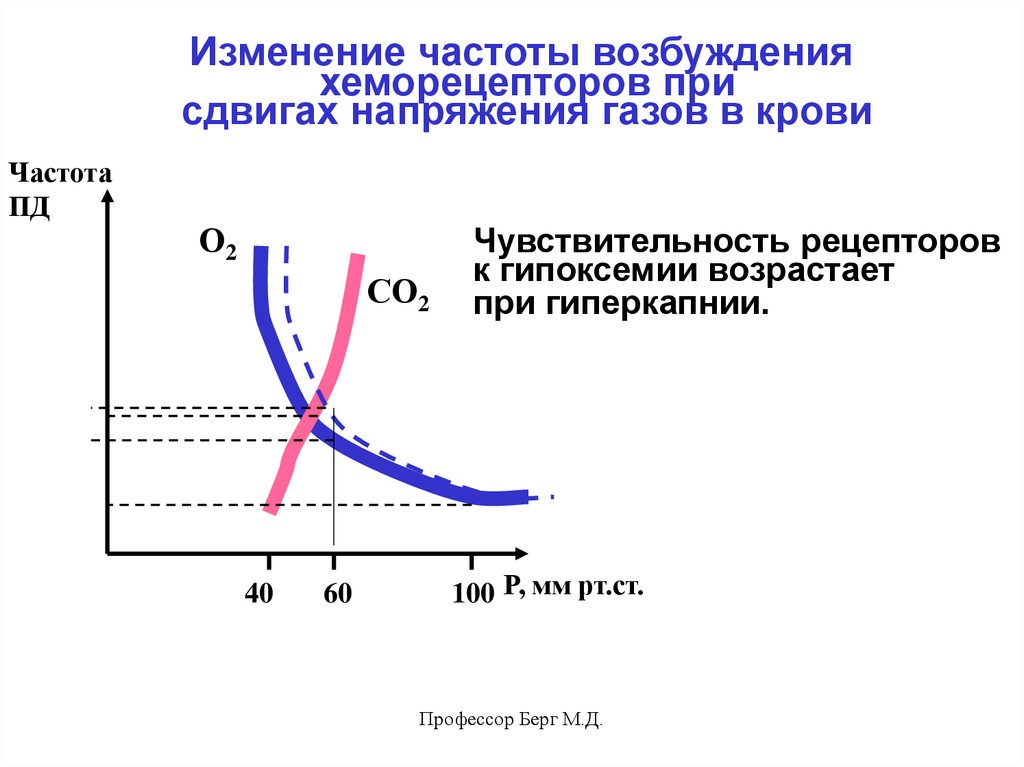
52.
Изменение частоты возбужденияхеморецепторов при
сдвигах напряжения газов в крови
Частота
ПД
О2
СО2
40
60
Чувствительность рецепторов
к гипоксемии возрастает
при гиперкапнии.
100 Р, мм рт.ст.
Профессор Берг М.Д.
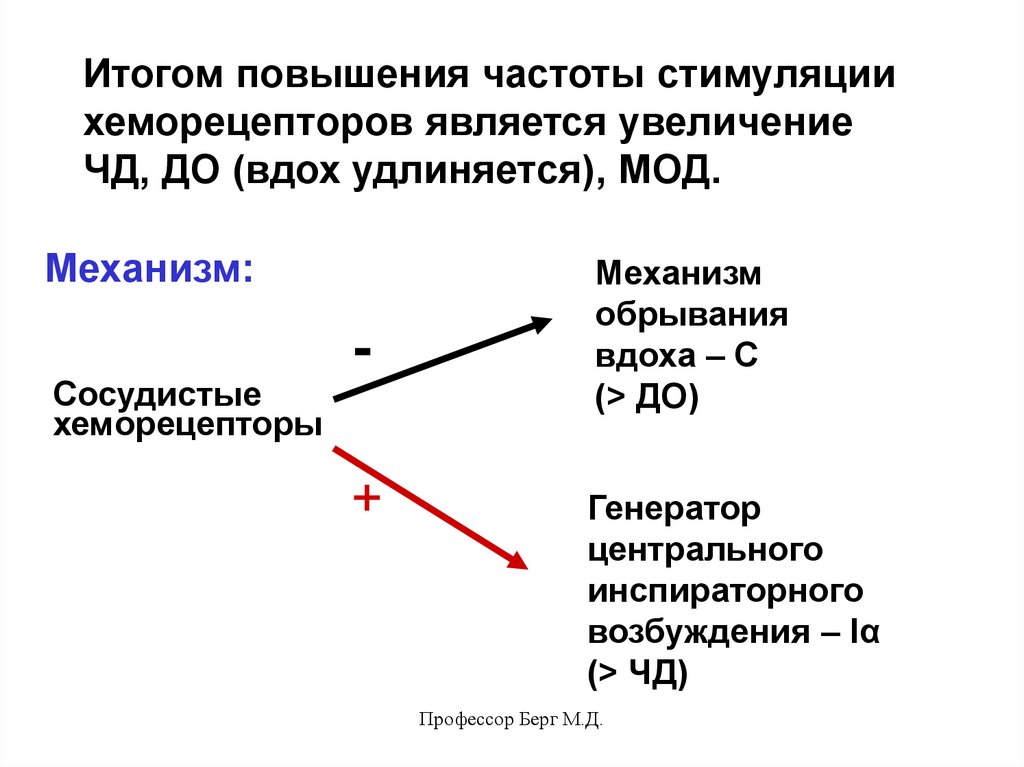
53.
Итогом повышения частоты стимуляциихеморецепторов является увеличение
ЧД, ДО (вдох удлиняется), МОД.
Механизм:
Сосудистые
хеморецепторы
+
Механизм
обрывания
вдоха – С
(> ДО)
Генератор
центрального
инспираторного
возбуждения – Iα
(> ЧД)
Профессор Берг М.Д.
54.
Тканевые хеморецепторы.Располагаются в межклеточном веществе
тканей, например метаболорецепторы
скелетных мышц.
Они чувствительны к сдвигам рН.
Сдвиг происходит при накоплении в
тканях СО2, молочной кислоты.
Профессор Берг М.Д.
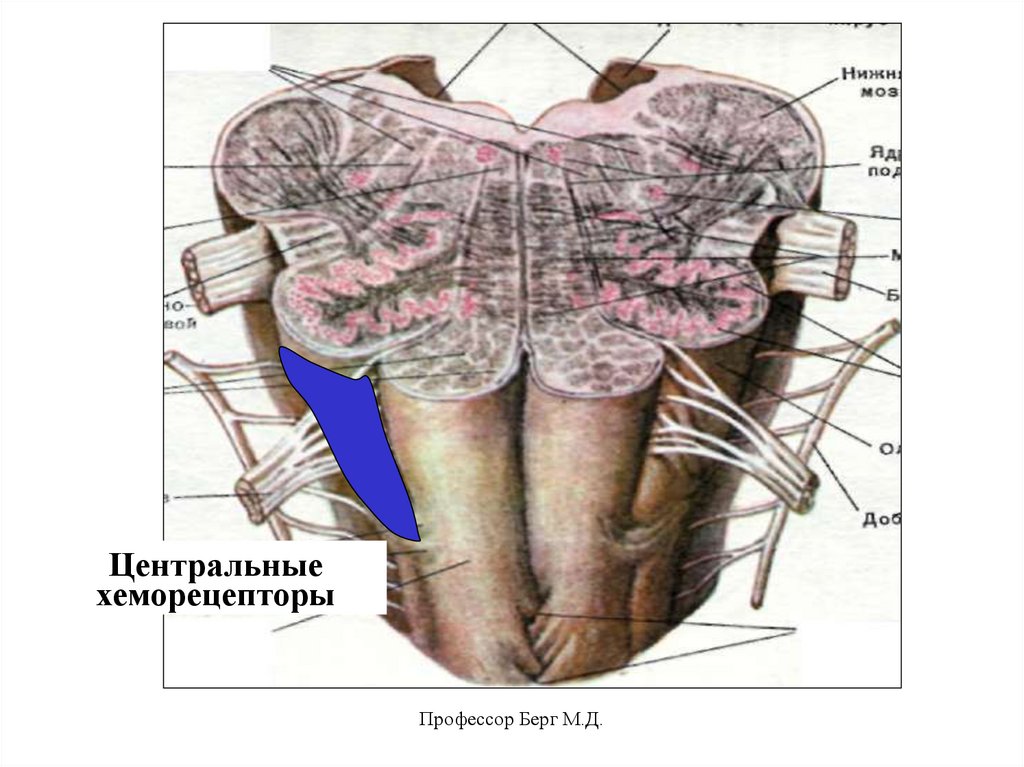
55.
Центральные хеморецепторы.Это нейроны, расположенные на переднебоковых поверхностях продолговатого
мозга, а также в некоторых других его
структурах.
Раздражители:
• увеличение Н+ в ликворе и межклеточной
жидкости,
• увеличение рСО2 в крови и ликворе
(спинно-мозговая жидкость).
Итогом является увеличение ЧД, ДО (вдох
удлиняется), МОД.
Профессор Берг М.Д.
56.
Центральныехеморецепторы
Профессор Берг М.Д.
57.
Рефлекторные влияния с рецепторовлегких, воздухоносных путей и
дыхательных мышц.
Профессор Берг М.Д.
58.
1. Рецепторы воздухоносных путей:• Механорецепторы носа. Вызывают
рефлекторное чихание.
• Механорецепторы носоглотки и глотки.
Управляют дыханием при глотании
(остановка дыхания, т.е. защита от попадания
пищи в дыхательные пути).
• Механорецепторы и хеморецепторы
гортани. Вызывают рефлекторную остановку
дыхания – апноэ, кашель
(с механорецепторов) для удаления
посторонних частиц.
Профессор Берг М.Д.
59.
• Механо- и хеморецепторы крупныхвоздухоносных путей (ирритантные
рецепторы) – рефлекторное апноэ, кашель,
сужение бронхов.
Рефлексы с этих рецепторов подразделяют на:
• Рефлексы, предохраняющие от попадания
чужеродных веществ (апноэ).
• Рефлексы, изгоняющие попавшие частицы
(кашель, чихание).
Чихание – форсированный выдох через нос
после открытия голосовой щели.
Кашель – форсированный выдох через рот
после открытия голосовой щели.
Профессор Берг М.Д.
60.
2. Механорецепторы легких.Участвуют в рефлексе Геринга-Брейера,
т.е. плавной смене вдоха на выдох.
3. Юкстаальвеолярные рецепторы
(J-рецепторы) расположены в межальвеолярных перегородках.
Являются хемо- и механорецепторами,
возбуждаются при воспалении, отеке легких,
при перерастяжении ткани легкого.
Вызывают бронхоспазм, частое и поверхностное дыхание (одышка), увеличение
секреции слизи.
Профессор Берг М.Д.
61.
4. Проприорецепторы дыхательныхмышц.
Любое препятствие к достижению
заданного ДЦ объема дыхания (ДО)
вызывает усиление импульсации от
рецепторов растяжения мышц и
рефлекторно усиливает их сокращение.
Профессор Берг М.Д.
62.
Функциональная система поддержанияпостоянства рО2 и рСО2
ПОВЕДЕНИЕ
ВНС
ЛРК
АУ
Центральные
хеморецепторы
ЖВС
Дыхательная
система
Система крови
Кровообращение
Почки
ГУМОРАЛЬНЫЙ ПУТЬ
НЕРВНЫЙ ПУТЬ
Профессор Берг М.Д.
рО2, рСО2
ХЕМОРЕЦЕПТОРЫ
63.
Дыхание плодаВ период внутриутробного развития формируется
ритмическая дыхательная активность.
Внутриутробные дыхательные движения начинаются
с 13-ой недели, а нормальный тип устанавливается с
27 по 40 недели (45-60 в мин, амплитуда – до 5 мм).
С началом родового акта дыхательные движения
плода затормаживаются. Послеродовое апноэ
является следствием общего угнетения активности
ЦНС плода.
Первый вдох происходит в результате возобновления
временно выключенной автоматии дыхательного
центра. Внеутробные дыхательные движения
начинаются не с вдоха, а с чихания (кашля), т.е
сильного рефлекторного движения, сопровождающегося выбросом слизи и прочего из носоглотки, носа
и ротовой полости, после чего возникает пауза, а
затем – дыхание. Его частота около 35 в мин, ДО –
15-25 мл.
Профессор Берг М.Д.
64.
Дыхание при разных состоянияхФизические нагрузки.
1. Внешнее дыхание.
Увеличение транспорта О2 и СО2 из внешней
среды: увеличение ДО (от 300-800 до 2000-3000
мл), увеличение ЧД от10-20 до 40-60 в мин.
МОД – от 5-6 л/мин до 170-300 л/мин.
2. Диффузия газов в легких.
Увеличение площади для диффузии с 60 до 120
м2 за счет глубины дыхания.
Профессор Берг М.Д.
65.
3. Транспорт газов кровью.• Увеличение количества эритроцитов и гемоглобина за счет рефлекторного сокращения
селезенки и выброса эритроцитарной массы
(до 500 мл).
• Увеличение скорости транспорта за счет повышения линейной скорости кровотока (см/с) в
2-3 раза.
• Увеличение объемного кровотока (мл/мин)
с 5-6 л/мин до 30 л/мин.
Профессор Берг М.Д.
66.
4. Диффузия газов в тканях• Увеличение диффузионной площади за счет
повышения числа функционирующих капилляров с 25-40% до 80-100%.
• Снижение диффузионного расстояния за счет
повышения числа функционирующих капилляров.
• Увеличение скорости диссоциации оксигемоглобина за счет повышения температуры в
работающих мышцах, закисления среды и повышения напряжения СО2.
Профессор Берг М.Д.
67.
Последствия физических тренировокУвеличение резервов транспорта на всех этапах.
1. Первый этап дыхания.
• Увеличение ЖЕЛ за счет объема грудной клетки, количества и силы мышечных волокон в
дыхательных мышцах.
• Снижение чувствительности центральных и
периферических хеморецепторов к гипоксии и
гиперкапнии. В покое – более редкое и глубокое
дыхание. Увеличивается время задержки
дыхания, необходимое, например при плавании
и нырянии.
Профессор Берг М.Д.
68.
2. Второй этап дыхания.• Увеличение площади альвеолярно-капиллярной
мембраны в покое и при нагрузках,
• в том числе за счет снижения физиологического
мертвого пространства.
3. Третий этап дыхания.
• Урежение ЧСС и увеличение УО в покое,
увеличение резерва ЧСС и резерва МОК.
• Повышение движущей силы кровотока за счет
скорости и силы сокращения сердца.
• Увеличение количества эритроцитов и гемоглобина.
• Повышение сродства гемоглобина к О2 (2,3-ДФГ).
Профессор Берг М.Д.
69.
• Улучшение перераспределительных реакцийкровообращения.
4. Четвертый этап дыхания.
• Повышение капилляризации мышц и сердца.
• Уменьшение диффузионного пути в тканях.
• Повышение концентрации миоглобина в
мышцах и сердце.
5. Улучшение нервной и гуморальной
регуляции на всех этапах транспорта газов.
Профессор Берг М.Д.
70.
Мерцательный эпителийПрофессор Берг М.Д.