


















")









")























")




 biology
biologySimilar presentations:






 функций в организме. Нервные центры. Возрастные особенности")
: торможение")


Свойства нервных центров, торможение в ЦНС, координационные принципы
1.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ,ТОРМОЖЕНИЕ в ЦНС,
КООРДИНАЦИОННЫЕ ПРИНЦИПЫ
2. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
1. Одностороннее проведение возбуждения.2. Центральная задержка рефлекса.
3. Последействие (кратковременное и реверберация)
4. Суммация: пространственная и временная. Центральное
облегчение и окклюзия.
5. Трансформация ритма.
6. Фоновая электрическая активность.
7. Тонус нервного центра (открыт Бронжестом) .
8. Утомление нервных центров.
9. Высокая чувствительность к гипоксии и глюкозе .
10. Посттетаническое усиление.
11. Пластичность (пресинаптическая и постсинаптическая;
гомосинаптическая и гетеросинаптическая).
3. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
1. одностороннее проведение возбуждения(химические синапсы),
2. центральная задержка проведения
возбуждения (химические синапсы),
4.
3. Последействие :1. кратковременное - связано с особенностью
синаптической передачи (медиатор не сразу
прекращает действовать, а лишь через время
(около 10 мс))
2. длительное - связано с циркуляцией
возбуждения по сетям
Лоренто де Но
5. 4. Временная и пространственная суммация
6. Центральное облегчение: при одновременном раздражении двух афферентов с перекрывающимися подпороговыми зонами количество
возбужденных нейронов больше, чем арифметическаясумма возбужденных нейронов при их раздельном
раздражении
А
1
При раздражении А
возбуждаются 2
нейрона (1,2)
2
3
При раздражении В
возбуждаются 2
нейрона (5, 6)
В
4
При раздражении А + В
возбуждаются 6 нейронов
(1, 2, 3, 4, 5, 6)
5
6
7.
Центральная окклюзия:при одновременном раздражении двух афферентов с
перекрывающимися пороговыми зонами количество
возбужденных нейронов меньше, чем арифметическая
сумма возбужденных нейронов при их раздельном
раздражении.
А
В
При раздражении А
возбуждаются 4
нейрона (1,2,3,4)
1
2
3
4
5
При раздражении А + В возбуждаются
6 нейронов (1, 2, 3, 4, 5, 6)
При раздражении В
возбуждаются 4
нейрона (3, 4, 5, 6)
6
8.
5. усвоение и трансформация ритма1. усвоение необходимо для ритмичной работы
НЦ (например, для ритмических движений).
2. трансформация ритма может быть
повышающей и понижающей, что связано с
особенностями синапсов данного НЦ
Порог 30 мв
Порог 10 мв
Тело нейрона
Ек
Ео
Аксонный холмик
Ек
Ео
9.
6. фоновая активность обеспечивается:поступлением импульсов от соседних НЦ
(дивергенция, конвергенция);
суммацией миниатюрных потенциалов,
возникающих за счет утечки медиатора
7. тонус НЦ необходим для обеспечения
готовности к рефлекторной деятельности
и возникает:
1. за счет спонтанной активности нейронов НЦ,
2. из-за постоянного поступления импульсации от
рецепторов с переферии
10. Опыт Бронжеста
Тонус нервного центаОпыт Бронжеста
11.
8. утомление в НЦ возникает из-за:1.
2.
3.
4.
истощения медиатора,
истощения АТФ и глюкозы,
снижения чувствительности рецепторов,
метаболических сдвигов в нейронах.
9. Высокая чувствительность к О2
(необратимые последствия: кора – 5-6
минут, ствол – 15-20 минут, спинной мозг –
30 минут) , глюкозе.
12.
10. Посттетаническая потенциацияЕсли , после интенсивного раздражения, дать синапсу отдохнуть и
нанести одиночное раздражение, то медиатора выбросится
значительно больше и ответная реакция возрастёт.
Это явление называется посттетанической потенциацией.
Причины:
- Во время тетанического раздражения усиливаются синтетические
процессы в нейронах, возрастает энергообмен.
- Увеличивается количество везикул, готовых к высвобождению.
Поэтому, обычное раздражение вызывает повышенный выброс
медиатора в синаптическую щель.
13.
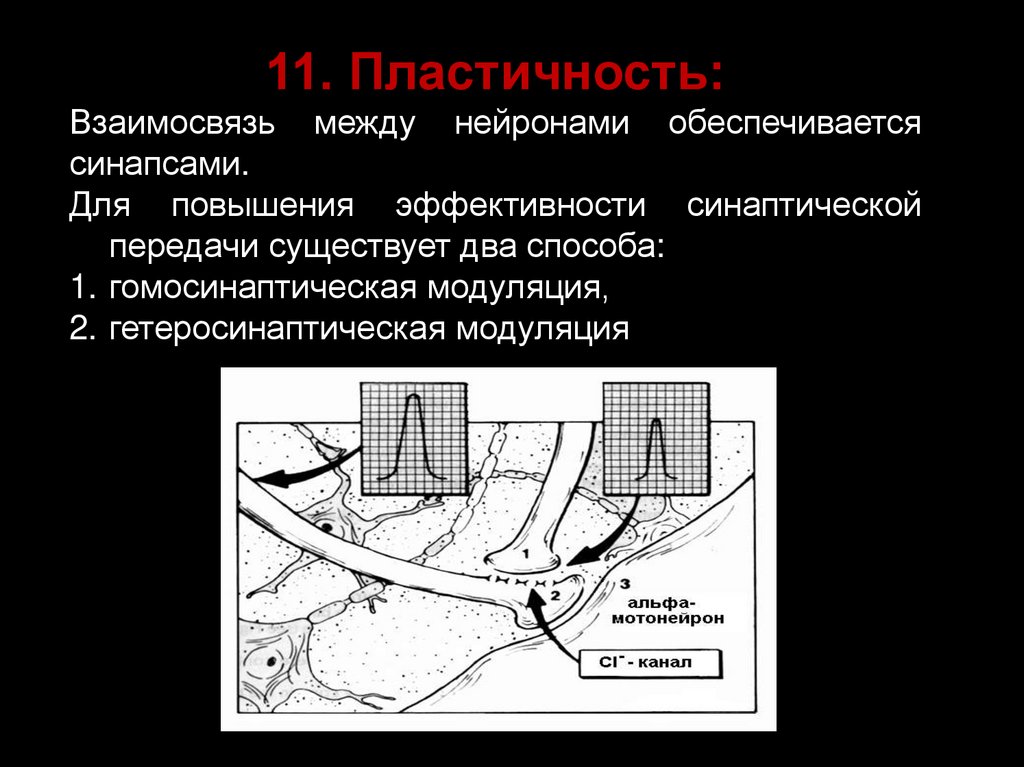
11. Пластичность:Взаимосвязь между нейронами обеспечивается
синапсами.
Для повышения эффективности синаптической
передачи существует два способа:
1. гомосинаптическая модуляция,
2. гетеросинаптическая модуляция
14. Гомосинаптическая модуляция
частое (тетаническое) раздражениепресинаптической мембраны
↓
повышение [Са2+ ] в синаптическом окончании
↓
увеличение выделения медиатора
↓
увеличение амплитуды ПСП
Это явление аналогично тетанической
потенциации.
15. Гомосинаптическая модуляция
• На постсинаптической мембране возрастаетконцентрация рецепторов, поэтому
повышается чувствительность к медиатору и,
как следствие, снижается время синаптической
задержки.
16. Гетеросинаптическая модуляция
Этапы:Выделение медиатора
→↑ цАМФ
→ инактивация S- К+-каналов
→ удлинение фазы
деполяризации ПД
→ больше входящего Са2+
→ больше выделение
медиатора
→ больше амплитуда ПСП
17. Торможение и его функции
• ТОРМОЖЕНИЕ - это самостоятельный нервный процесс,вызываемый возбуждением и проявляющийся в подавлении
другого возбуждения.
В отличие от возбуждения, торможение может развиваться
только в форме локального ответа.
• ФУНКЦИИ ТОРМОЖЕНИЯ:
• 1. Координация рефлекторных реакций;
• 2. Охранительное, т.е. защита нервных центров от истощения
и утомления;
• 3. Участие в переработке информации;
• 4. Ограничение возбудительного процесса;
• 5. Формирование условных рефлексов.
18. Виды торможения
1. ЦЕНТРАЛЬНОЕ (Сеченовское)2. ВОЗВРАТНОЕ
а) собственно возвратное
б) латеральное
в) медиальная зона
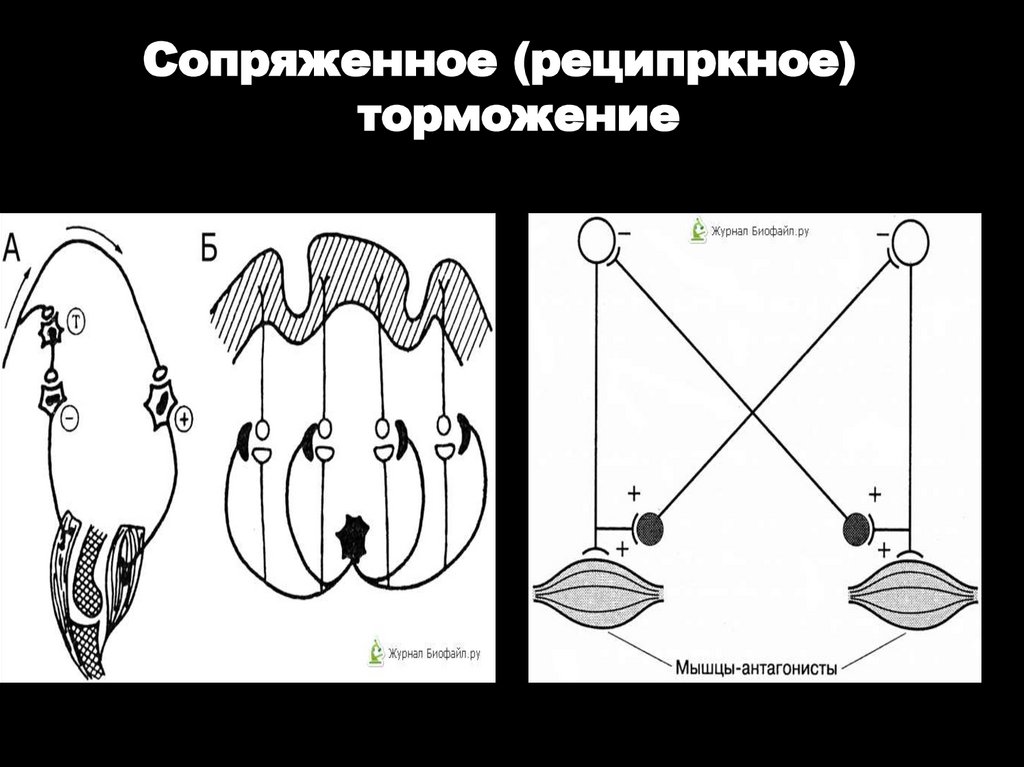
3. РЕЦИПРОКНОЕ
4. ТОРМОЖЕНИЕ ВСЛЕД ЗА ВОЗБУЖДЕНИЕМ
5. ПЕССИМАЛЬНОЕ (по Н.Введенскому)
19. Тормозные нейроны
В ЦНС имются нейроны со специфической функцией торможения :
клетки Реншоу спинного мозга, клетки Пуркинье мозжечка,
корзинчатые клетки гиппокампа, входящего в состав лимбической
системы и др.
В коре головного мозга 4 вида тормозных клеток:
большие корзинчатые нейроны - 3, 4, 5 слои коры головного
мозга, их аксоны сильно ветвятся и образуют сплетения на
площади около 500 мкм. Они тормозят активность нейронов 3, 4, 5
слоев;
малые корзинчатые клетки нейроны - 2, 3 слои коры - их
аксоны ветвятся на меньшей площади, около 50 мкм, и тормозят 2
и 3 слои;
нейроны с кистеобразным аксоном - 1 слой коры,
образует аксон, на конце которого разветвление в виде кисти;
тормозит клетки 1-го слоя;
канделяброобразные нейроны - на границе между 2 и 3
слоями, вниз от них идет аксон и дает несколько ответвлений
вверх, тормозят активность всех слоев.
20. ТИПЫ ТОРМОЖЕНИЯ
• П Е Р В И Ч Н О Е:А) ЦЕНТРАЛЬНОЕ (СЕЧЕНОВСКОЕ)
Б) ВОЗВРАТНОЕ
В) РЕЦИПРОКНОЕ
• В Т О Р И Ч Н О Е:
А) ПЕССИМАЛЬНОЕ (по Н.Введенскому)
Б) ТОРМОЖЕНИЕ ВСЛЕД ЗА ВОЗБУЖДЕНИЕМ
21. Торможение в ЦНС (опыт И.М. Сеченова)
22.
23.
24. Окклюзия
А1
При раздражении А
возбуждаются 4
нейрона (1,2,3,4)
2
3
При раздражении В
возбуждаются 4
нейрона (3, 4, 5, 6)
В
4
5
6
Клетки
центральной
части
нейронного пула
При раздражении А + В
возбуждаются 6
нейронов (1, 2, 3, 4, 5, 6)
25. ТОРМОЖЕНИЕ В ЦНС
а) Собственно возвратное торможение (по Реншоу)б) Латеральное торможение
26.
Медиальная тормозная зона27.
Сопряженное (реципркное)торможение
28. Торможение вслед за возбуждением
• Суммация следовой гиперполяризациипосле серии возбуждений
29. Пессимальное торможение
1. При частом раздражении постсинаптические потенциалы
суммируются,
что
приводит
к
стойкой
деполяризации
постсинаптической мембраны (это является причиной десинтезации)
и развитию блока проведения.
2. При длительном раздражении нерва нарушение синаптической
передачи может развиваться в связи с истощением запасов
медиатора. Кроме этого, под влиянием продуктов обмена в
межсинаптической
щели
может
происходить
снижение
чувствительности к АЦХ.
3. Когда частота стимулов очень высокая, проведение возбуждения с
нерва на мышцу может быть блокировано на пресинаптической
мембране (она безмиелиновая), которая обладает меньшей
лабильностью, чем миелиновая мембрана нервного волокна.
Данный механизм нарушения передачи возбуждения сходен с
катодической депресией.
30. Механизмы торможения:
I. Пресинаптическое торможениеII. Постсинаптическое торможение
31. Механизм постсинаптического торможения
• Связан с воздействием тормозного медиатора (глицина) напостсинаптическую мембрану и развитием ТПСП, как следствие
– снижением возбудимости.
• Глицин увеличивает выход ионов K+ из клетки и вход ионов Cl– в клетку через постсинаптическую мембрану. За счет
электротонического распространения происходит увеличение
МП. Глицин является основным тормозным медиатором
спинного мозга.
• ТПСП – представляет собой зеркальное отражение ВПСП с
временем нарастания 1-2 мс и спада 10-12 мс.
• Известно 2 яда, которые блокируют передачу в тормозных
синапсах мотонейронов
спинного мозга, вызывая судороги:
стрихнин и столбнячный токсин.
32. Постсинаптическое торможение Тормозной постсинаптический потенциал ( ТПСП )
мвЕк
Ео
- 70
- 74
ГИПЕРПОЛЯРИЗАЦИЯ
33.
34.
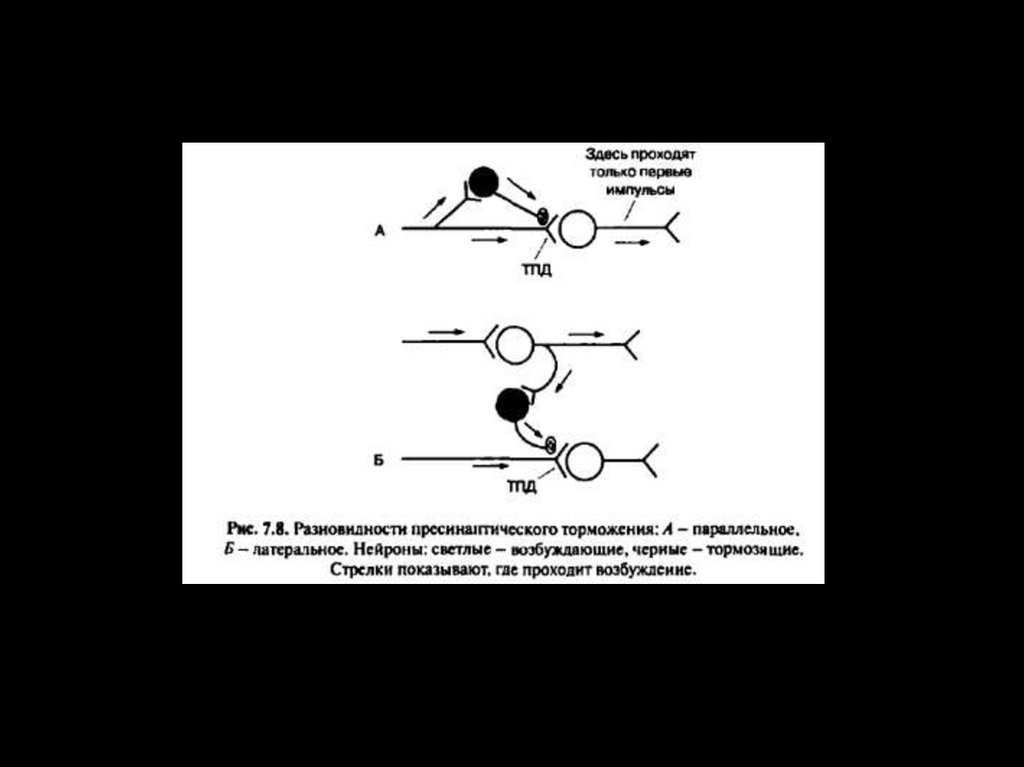
35. МЕХАНИЗМ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ
Возникает в области аксо-аксональных синапсов, на пресинаптических
терминалях. Сопровождается медленной и длительной деполяризацией
пресинаптической мембраны, вследствие активации ГАМК выхода ионов Clнаружу.
При этом происходит инактивация Na+-каналов, уменьшение амплитуды ПД,
уменьшение
входящего
Ca2+
тока
и
уменьшение
количества
высвобождающегося медиатора.
Временной ход пресинаптического торможения более длительный по
сравнению с постсинаптическим торможением – время нарастания 15-20 мс,
спада – 100/150 мс. Возникает, так называемая, деполяризация первичных
афферентов.
36.
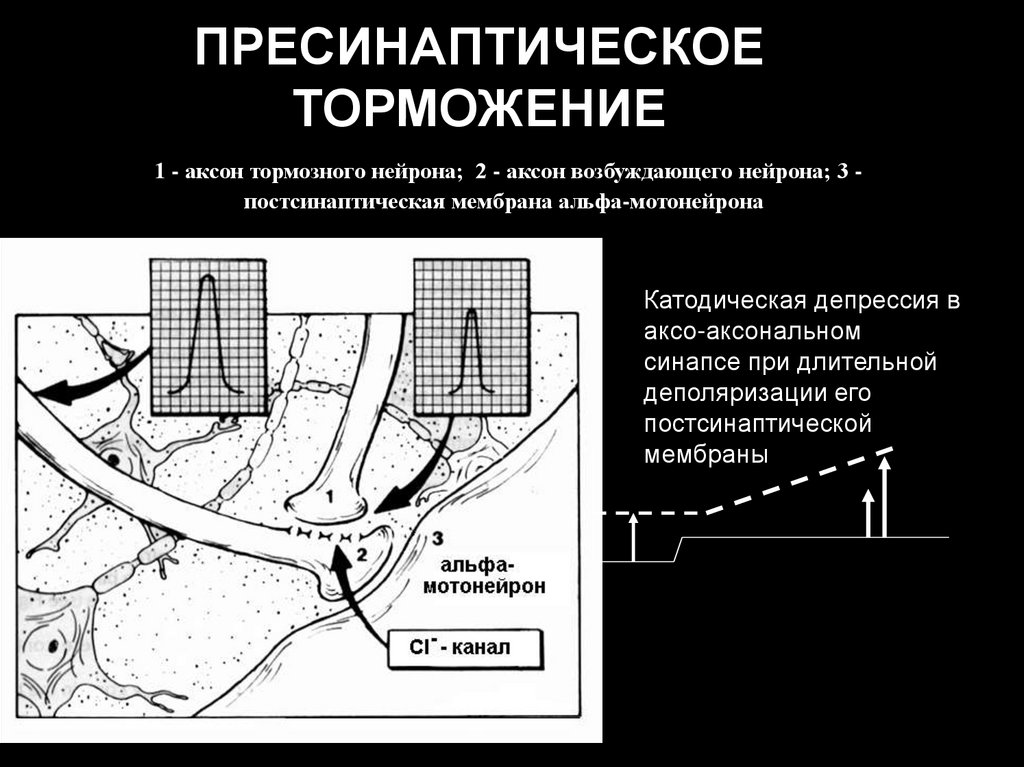
ПРЕСИНАПТИЧЕСКОЕТОРМОЖЕНИЕ
1 - аксон тормозного нейрона; 2 - аксон возбуждающего нейрона; 3 постсинаптическая мембрана альфа-мотонейрона
Катодическая депрессия в
аксо-аксональном
синапсе при длительной
деполяризации его
постсинаптической
мембраны
37.
38.
39.
Антиноцицептивная система выделяет биологическиактивные эндогенные опиоидные вещества – это «внутренние
наркотики».
Они называются эндорфины, энкефалины, динорфины. Все
они
по
химическому
строению
являются
короткими пептидными цепочками, как бы кусочками белковых
молекул, т. е. состоят из аминокислот.
Отсюда и название: нейропептиды, опиоидные пептиды.
Опиоидные — т. е. подобные по действию наркотическим
веществам опийного мака.
На
многих
нейронах
болевой
системы
существуют
специальные молекулярные рецепторы к этим веществам. Когда
опиоиды связываются с этими рецепторами, то возникает
пресинаптическое и/или постсинаптическое торможение в
нейронах болевой системы.
Болевая ноцицептивная система тормозится и слабо
реагирует на боль.
На рисунке более мелкий АНЦ-нейрон (он слева) тормозит
синапс болевого нейрона и мешает ему передавать болевое
возбуждение дальше.
Налоксо́н — антагонист опиоидных рецепторов, применяется
как антидот при передозировках опиоидов.
40.
41. Примеры нарушения торможения в ЦНС
• НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ:• СТРИХНИН
–
КОНКУРИРУЕТ
С
ГЛИЦИНОМ ЗА
СВЯЗЫВАНИЕ С РЕЦЕПТОРАМИ НА ПОСТСИНАПТИЧЕСОЙ
МЕМБРАНЕ
• СТОЛБНЯЧНЫЙ ТОКСИН - НАРУШАЕТ ОСВОБОЖДЕНИЯ
ГЛИЦИНА ИЗ ПРЕСИНАПТИЧЕСКИХ ОКОНЧАНИЙ
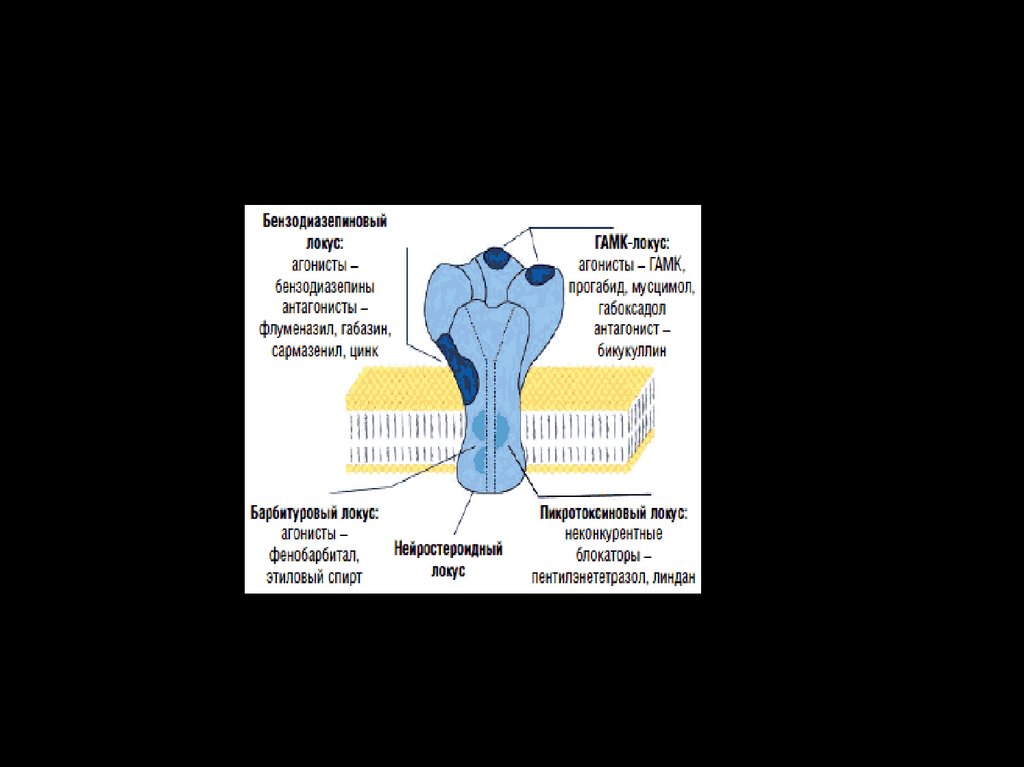
42. Блокаторы и агонисты ГАМК-рецепторов
Блокаторы и агонисты ГАМКрецепторовОсновным ядом, блокирующим пресинаптическое торможение является
бикукулин.
Вещества, усиливающие ингибиторный эффект ГАМК, - релаксанты и
транквилизаторы.
Из яда мухомора получают агонист ГАМК-рецепторов – мусцимол,
который применяется для блокады судорог.
43.
Растения семейства дымянковых Fumariaceae (хохлаткаполая, хохлатка Горчакова, хохлатка Маршалла) содержат
ядовитые алкалоиды группы бульбокапнина - бульбокапнин,
бикукулин и др.
44. Координация нервных процессов
Обеспечивает согласованную
деятельность всех органов организма и
адекватную реакцию на различные
раздражители .
45. Функции координированной деятельности:
1) обеспечивает четкое выполнениеопределенных функций, рефлексов;
2) обеспечивает последовательное включение в
работу различных нервных центров для
обеспечения сложных форм деятельности;
3) обеспечивает согласованную работу
различных нервных центров.
46. ПРИНЦИПЫ КООРДИНАЦИИ РЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ В ЦНС
1. КОНВЕРГЕНЦИИ ИЛИ ОБЩЕГО КОНЕЧНОГО ПУТИ (по Шеррингтону)
2. ДИВЕРГЕНТНОСТИ (ИРРАДИАЦИЯ)
3. ОБРАТНОЙ АФФЕРЕНТАЦИИ
4. ДОМИНАНТА
5. СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
6. РЕЦИПРОКНОСТИ
7. СОДРУЖЕСТВЕННОЙ РАБОТЫ НЕРВНЫХ ЦЕНРОВ
8. ПЛАСТИЧНОСТИ НЕРВНЫХ ЦЕНТРОВ
47.
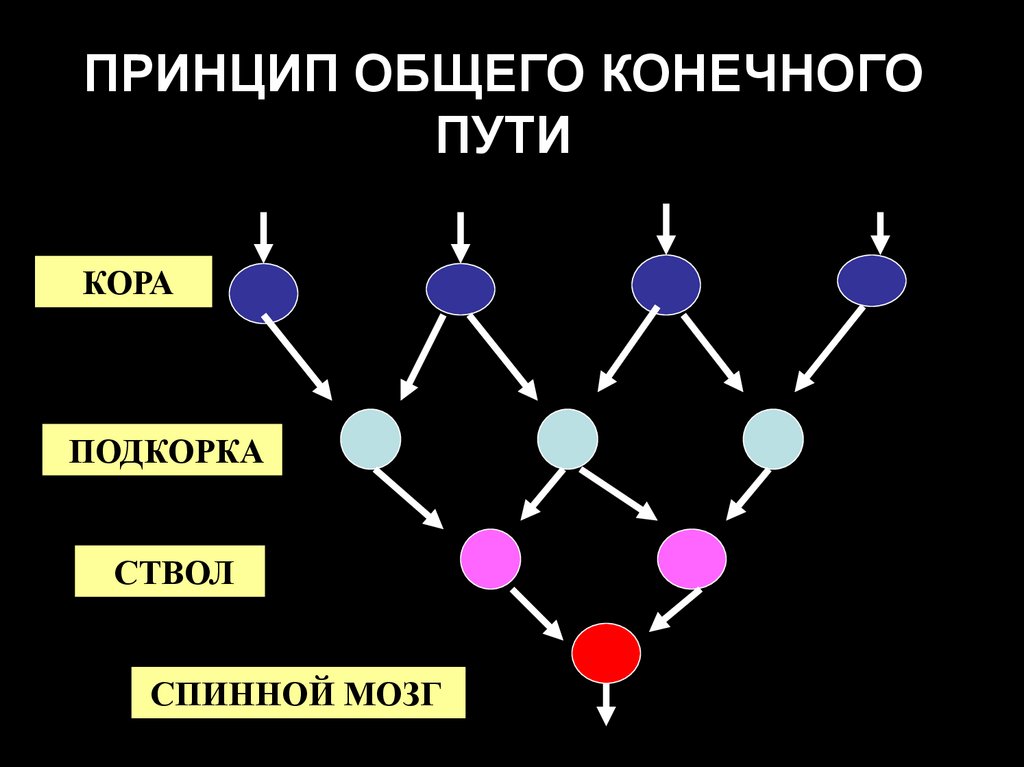
ПРИНЦИП ОБЩЕГО КОНЕЧНОГОПУТИ
КОРА
ПОДКОРКА
СТВОЛ
СПИННОЙ МОЗГ
48. Принцип общего конечного пути в спинном мозге
49.
50.
РАСПРОСТРАНЕНИЕ НЕРВНЫХ ИМПУЛЬСОВ В ЦНСДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
ДИВЕРГЕНЦИЯ НЕРВНЫХ ИМПУЛЬСОВ В ЦНС
Рецептор
51.
52.
53.
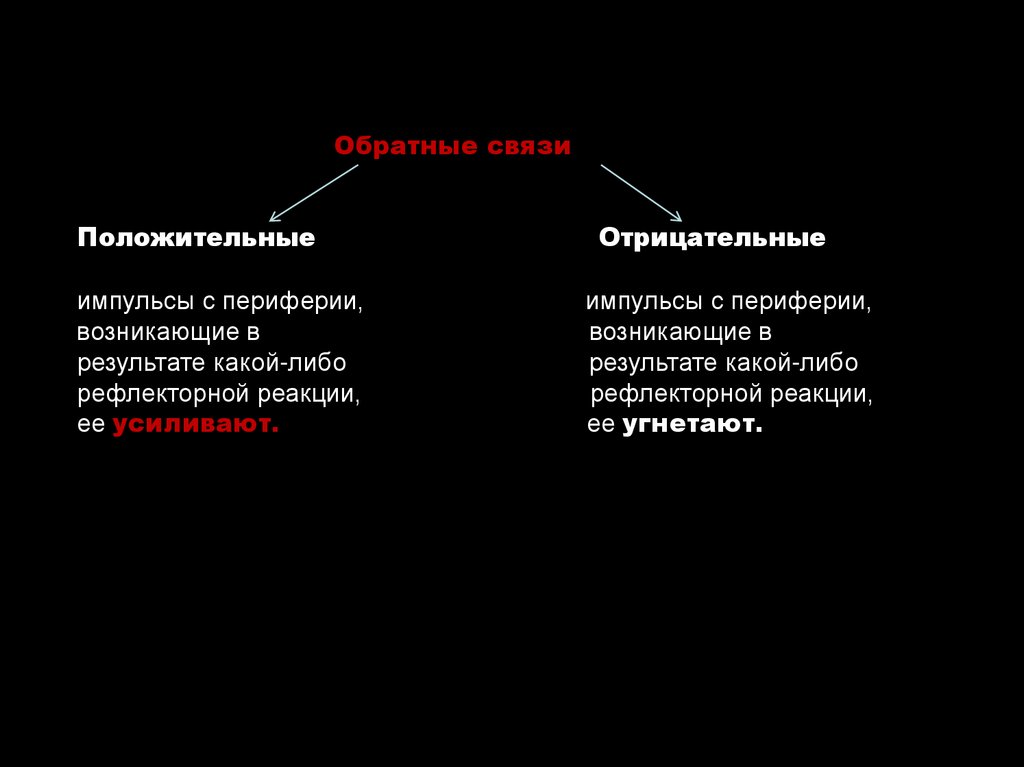
Обратные связиПоложительные
импульсы с периферии,
возникающие в
результате какой-либо
рефлекторной реакции,
ее усиливают.
Отрицательные
импульсы с периферии,
возникающие в
результате какой-либо
рефлекторной реакции,
ее угнетают.
54.
РЕФЛЕКТОРНАЯ ДУГА И РЕФЛЕКТОРНОЕ КОЛЬЦОНЕРВНЫЙ ЦЕНТР
АФФЕРЕНТНЫЙ ПРОВОДНИК
РЕЦЕПТОР
РАЗДРАЖИТЕЛЬ
ЭФФЕРЕНТНЫЙ ПРОВОДНИК
ОБРАТНАЯ
СВЯЗЬ
ЭФФЕКТОР
55. ПРИНЦИП ОБРАТНОЙ АФФЕРЕНТАЦИИ
56.
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ( ПО А.А.Ухтомскому, 1931)
Доминанта
временно
господствующий
рефлекс
или
поведенческий
акт,
которым трансформируется
и направляется для данного
времени при прочих равных
условиях
работа
прочих
рефлекторных
дуг,
рефлекторного аппарата и
поведения в целом
57.
Основные cвойства доминанты( по А.А.Ухтомскому)
1. Повышенная возбудимость доминантного центра
2. Способность к суммации (подкреплять свое
возбуждение посторонними импульсами)
3. Стойкость возбуждения в доминантном центре
(способность тормозить другие текущие рефлексы
на общем конечном пути)
4. Инертность доминантного центра
58. Виды доминант
• Внутренние доминанты – ведущуюроль играет гормональный фон.
• Внешние доминанты – ведущую роль
играет психическая деятельность.
59. Причины прекращения существования доминанты
1.Достижение результата2.Появление новой, более значимой
доминанты
3.Запредельное торможение
60. ПРИНЦИП СУБОРДИНАЦИИ НЕРВНЫХ ЦЕНТРОВ
61.
62. ПРИНЦИП РЕЦИПРОКНОСТИ ( СОПРЯЖЕННОГО ТОРМОЖЕНИЯ )
++
+
-
-
63. Динамика основных нервных процессов:
Процессы иррадиации и концентрации составляют основу
индукционных отношений в ЦНС.
Индукция – свойство возбуждения или торможения вызывать
вокруг себя или после себя противоположный эффект.
Если очаг возбуждения или торможения вызывает в рядом
расположенном участке коры противоположный эффект – это
называется одновременной индукцией.
Если после прекращения возбуждения или торможения в данном
участке коры возникает противоположный эффект – это
последовательная индукция.
64.
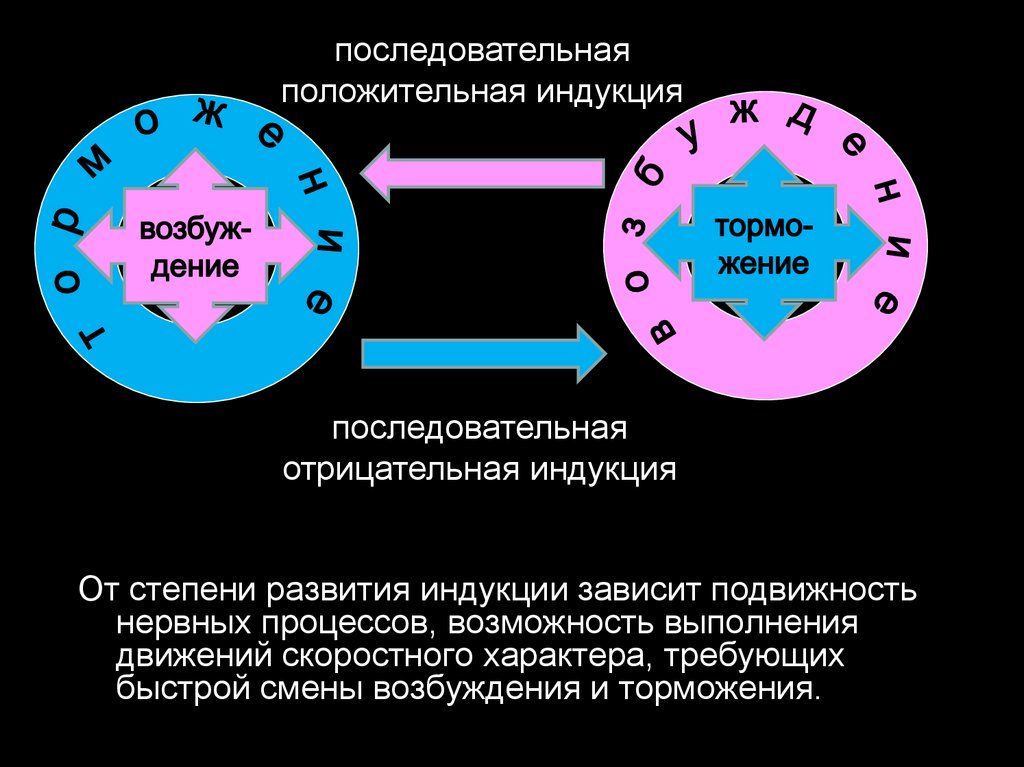
Возникновение торможения при последовательной индукции
называется отрицательной последовательной индукцией.
Возникновение возбуждения при последовательной индукции
называется –
положительной последовательной индукцией.
Одновременное возникновение возбуждения
торможения называется –
одновременной положительной индукцией.
вокруг
очага
Одновременное возникновение участка торможения, вокруг очага
возбуждения называется –
одновременная отрицательная индукция.
65.
9. Принцип индукцииИндукция ограничивает распространение
нервных процессов и обеспечивает
концентрацию.
одновременная
отрицательная
индукция
одновременная
положительная
индукция
66.
последовательнаяположительная индукция
последовательная
отрицательная индукция
От степени развития индукции зависит подвижность
нервных процессов, возможность выполнения
движений скоростного характера, требующих
быстрой смены возбуждения и торможения.