


")






















 biology
biologySimilar presentations:


")







Физиология мышц
1. Физиология мышц
1.2.
3.
4.
5.
Проведение ПД по аксону мотонейрона
Передача ПД через синапс
Скелетные (поперечно-полосатые) мышцы.
Шаговый механизм мышечных
сокращений.
Гладкие мышцы.
2. Скелетные мышцы начинают сокращаться лишь после того как получат ПД от мотонейрона. Проведение ПД от мотонейрона по миелинизированному ак
Скелетные мышцы начинают сокращаться лишьпосле того как получат ПД от мотонейрона.
Проведение ПД от мотонейрона по миелинизированному
аксону: ПД проводится сальтоторно – от одного
перехвата Ранвье к следующему. Это ускоряет
передачу ПД (до 100 м/с).
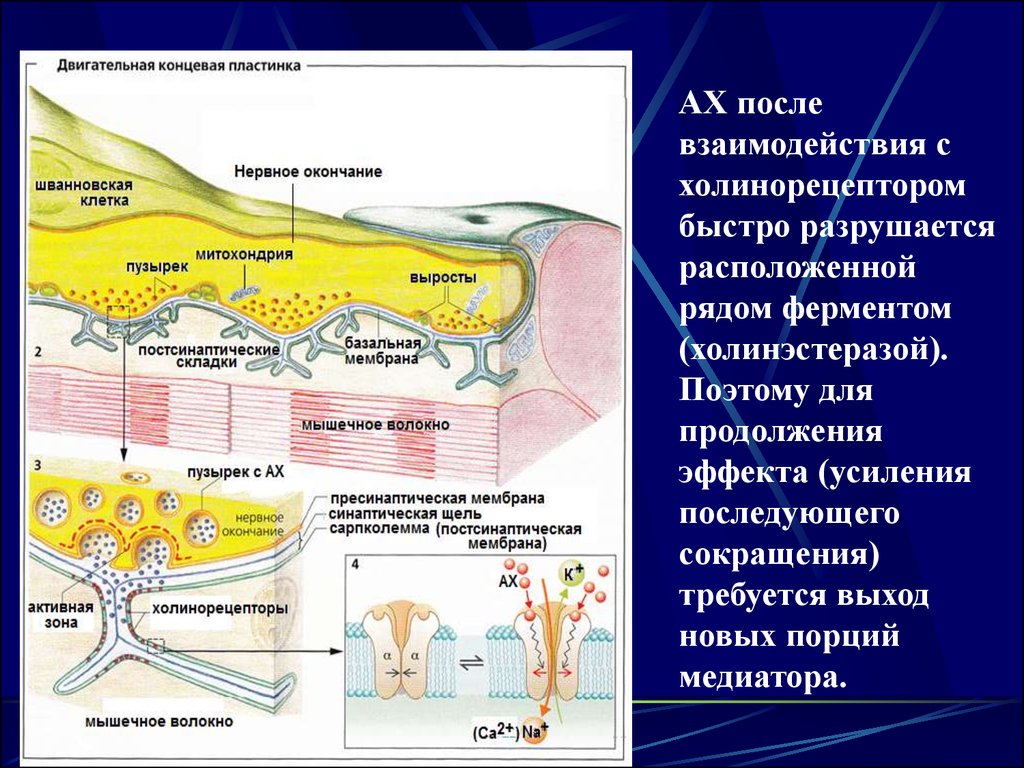
3. Нервно-мышечный синапс
Синаптическая щель:Пространство между двумя мембранами
Ширина - 20-30 нм
Заполнена синаптической жидкостью, своим
составом напоминающей плазму крови.
4. Медиатор (химический посредник - лиганд)
Внутри нервного окончания имеется большоеколичество (до 300.000) синаптических
пузырьков (диаметром около 50 нм), содержащих
химическое соединение ацетилхолин (АХ).
Это химический передатчик возбуждения, носящий
название - медиатор.
Каждый пузырек содержит «квант» медиатора около 104 молекул АХ.
В синаптической бляшке (окончание нейрона)
содержится большое количество митохондрий, что
свидетельствует о высокой метаболической
активности данного отдела нервного волокна.
5. Выход медиатора в нервно-мышечном синапсе
При поступлении ПД кпресинаптической мембране в ней
открываются Са2+ каналы.
Кальций поступает внутрь
нервного окончания.
Здесь кальций взаимодействует с
белком кальмодулином.
В результате несколько
пузырьков с медиатором
подтягиваются к мембране.
Медиатор выделяется в
синаптическую щель и
диффундирует к
постсинаптической мембране сарколемме.
6. Взаимодействие медиатора с постсинаптической мембраной
Медиатор диффундирует по синаптической жидкости ибольшая часть его молекул достигает
постсинаптической мембраны, где взаимодейст-вует с
холинорецептором (ХР) – периферическим белком
расположенным рядом с каналами.
Результатом взаимодействия АХ с ХР является
открытие хемовозбудимых ионных каналов.
Селективный участок его имеет диаметр 0,65 нм. Через
него могут проходить лишь положительные ионы
(стенка канала электроотрицательна) натрия или
кальция. Но в норме превалирует поток ионов натрия.
Они по концентрационному градиенту из
синаптической щели поступают внутрь мышечного
волокна и деполяризуют постсинаптическую мембрану.
7. Действие медиатора
Выброс медиатораобеспечивает
взаимодействие его с
лигандзависимыми
структурами канала
(ХР).
При этом ворота
открываются и ионы
(преимущественно
натрий) поступает
внутрь.
8. Переход ПКП в ВПСП
ПД по нерву могут поступать с максимальнойчастотой до 1000 имп/с.
Обычно по нерву поступает несколько десятков
ПД друг за другом («пачка»).
В связи с тем, что рецепторы от предыдущего
кванта АХ освобождаются очень быстро (уже
через 1-1,5 мс он разрушается холинэстеразой),
то каждое новое выделение медиатора приводит
к повторному открытию ионных каналов.
Возникший новый ПКП наслаивается на еще не
исчезнувшую предыдущую деполяризацию,
суммируясь, увеличивает его амплитуду.
9.
АХ послевзаимодействия с
холинорецептором
быстро разрушается
расположенной
рядом ферментом
(холинэстеразой).
Поэтому для
продолжения
эффекта (усиления
последующего
сокращения)
требуется выход
новых порций
медиатора.
10. Явление суммации
Обычно для передачитолько одного ПД
высвобождается до
миллиона молекул АХ
(200-300 везикул).
При этом мембрана
деполяризуется на
-0,3 мВ.
Для достижения
критического уровня
необходима
деполяризация до –50
мВ.
Значит необходимо
поступление новых ПД
11. Восстановление медиатора в синаптической бляшке
В нервном волокне происходит постоянноепополнение медиатора. Здесь имеется несколько
механизмов восстановления везикул с медиатором.
медиатор разрушается под действием фермента холинэстеразы на холин и уксусную кислоту.
Большая часть продуктов гидролиза ацетилхолина
возвращается в синаптическую бляшку, где участвует в
ресинтезе новых молекул медиатора, который
поступает во вновь формирующиеся везикулы.
Вторым путем восстановления потраченного
медиатора являются активные процессы местного
синтеза АХ из других сырьевых источников с помощью
соответствующих ферментов, имеющихся в
пресинаптическом окончании.
Третий путь: «подвоз» медиатора от тела нейрона аксонный транспорт.
12. Проведение ПД по мембране НА рис я напминаю как ПД проводится по сарколемме от точки к каждой соседней ранее не возбужденной точке
13.
СПР – депо Са2+Под влиянием ПД, проводящемуся по мембране мышечного
волокна и заходящему на мембрану СПР, из СПР выходит
кальций (здесь открываются кальциевые каналы).
Когда концентрация Са2+ в саркоплазме возрастает в 100 и
более раз (с 10-7 до 10-4 М/л) – начинается мышечное
сокращение.
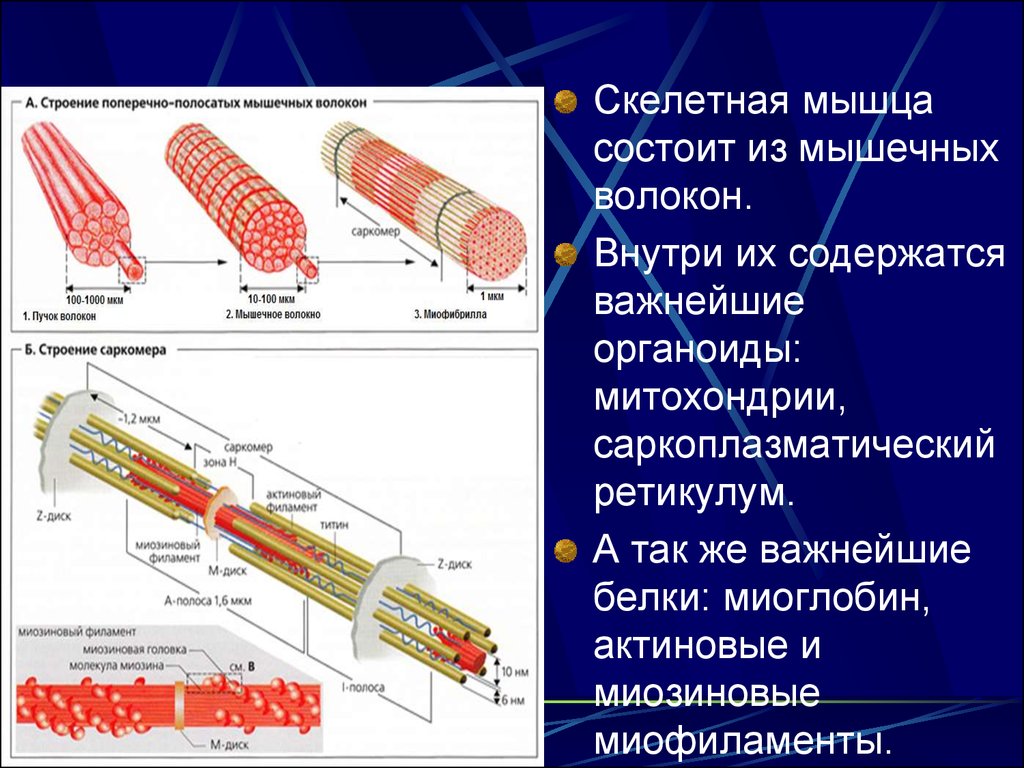
14.
Скелетная мышцасостоит из мышечных
волокон.
Внутри их содержатся
важнейшие
органоиды:
митохондрии,
саркоплазматический
ретикулум.
А так же важнейшие
белки: миоглобин,
актиновые и
миозиновые
миофиламенты.
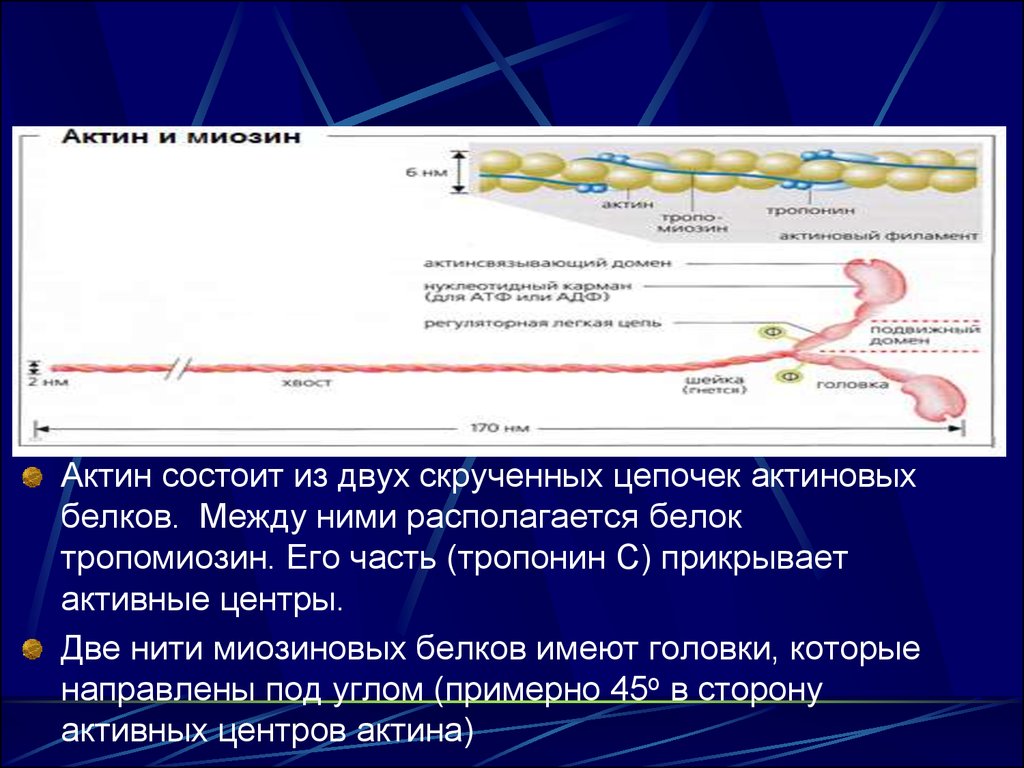
15.
Актин состоит из двух скрученных цепочек актиновыхбелков. Между ними располагается белок
тропомиозин. Его часть (тропонин С) прикрывает
активные центры.
Две нити миозиновых белков имеют головки, которые
направлены под углом (примерно 45о в сторону
активных центров актина)
16. Схема взаимодействия актиновых и миозиновых филаментов
17. Взаимодействие актиновых и миозиновых филаментов
10080
60
Восто
к
40
Запад
20
Север
0
1 2 3 4
кв кв кв кв
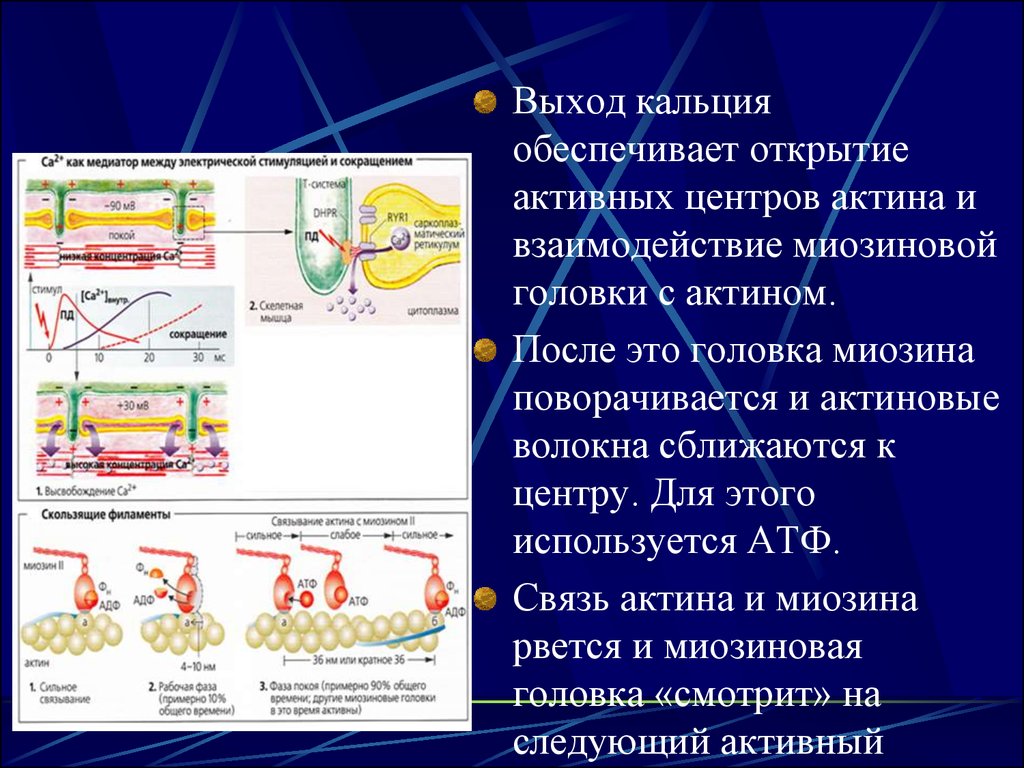
18.
Выход кальцияобеспечивает открытие
активных центров актина и
взаимодействие миозиновой
головки с актином.
После это головка миозина
поворачивается и актиновые
волокна сближаются к
центру. Для этого
используется АТФ.
Связь актина и миозина
рвется и миозиновая
головка «смотрит» на
следующий активный
19. Шаг
Указанное вышеименуется
«шаговым»
механизмом.
20. Этапы «шагового» механизма
Последовательные этапы:а – расслабление,
б – соединение миозиновых
головок с активным центром
актина,
в – поворот головки миозина и
сближение - мембран,
г – разрыв связи миозина с
актином.
21. Кальмодулин
- Тропонин С, имеющемуся в тонких нитях,идентичен кальмодулину.
Присоединяя Са2+, кальмодулин
способствует активации АТФазы и
использованию энергии АТФ для связи
активного центра актиновой нити и головки
миозина и последующему движению шейки
миозиновой головки (укорочению мышцы).
22. Кальций
Деполяризация мембраны цистерн открывает здесьэлектровозбудимые кальциевые каналы.
В связи с тем, что в саркоплазме концентрация
кальция менее 10-7 М/л, а в саркоплазматическом
ретикулуме - более 10-4 М/л, начинается интенсивный
выход ионов Са2+ в саркоплазму.
Выделившийся кальций и является инициатором
мышечного сокращения.
Достаточный для начала мышечного сокращения
уровень кальция (10-5 М/л) достигается через 12-15 мс
после прихода нервного импульса. Это скрытое,
латентное время мышечного сокращения.
В связи с тем, что скорость распространения ПД по
сарколемме выше времени, необходимого для
выделения Са2+ из саркоплазматического ретикулума,
то все фибриллы участка мышцы, иннервируемого
одним нервом, сокращаются одновременно.
23. Роль кальция в мышечном сокращении
1 – выброс медиатора в синаптическую щель.2 – освобождение активного центра актина.
3 – расслабление мышцы (разрыв связи миозина
с актином – кальциевая АТФ-аза).
24. Соотношение ПД и рефрактерности у различных мышц
Продолжительность периода рефрактерности ускелетной мышцы небольшая, поэтому при
поступлении по нерву частых ПД они могут не
расслабляясь сильнее и продолжительнее
сокращаться (см. далее).
25. Различные режимы сокращения скелетной мышцы
а - одиночное сокращение,б – неполный тетанус,
в – полный тетанус.
26. Роль АТФ в мышечном сокращении
а) сокращения (образования мостиков);б) расслабления (разрыва мостиков);
в) работы Са2+-насоса (2 АТФ на один ион Са2+);
г) работы Nа+,К+-насоса.
Однако в саркоплазме мышцы АТФ относительно немного. Ее
хватит лишь на несколько мышечных сокращений (примерно
8 одиночных сокращений).
27.
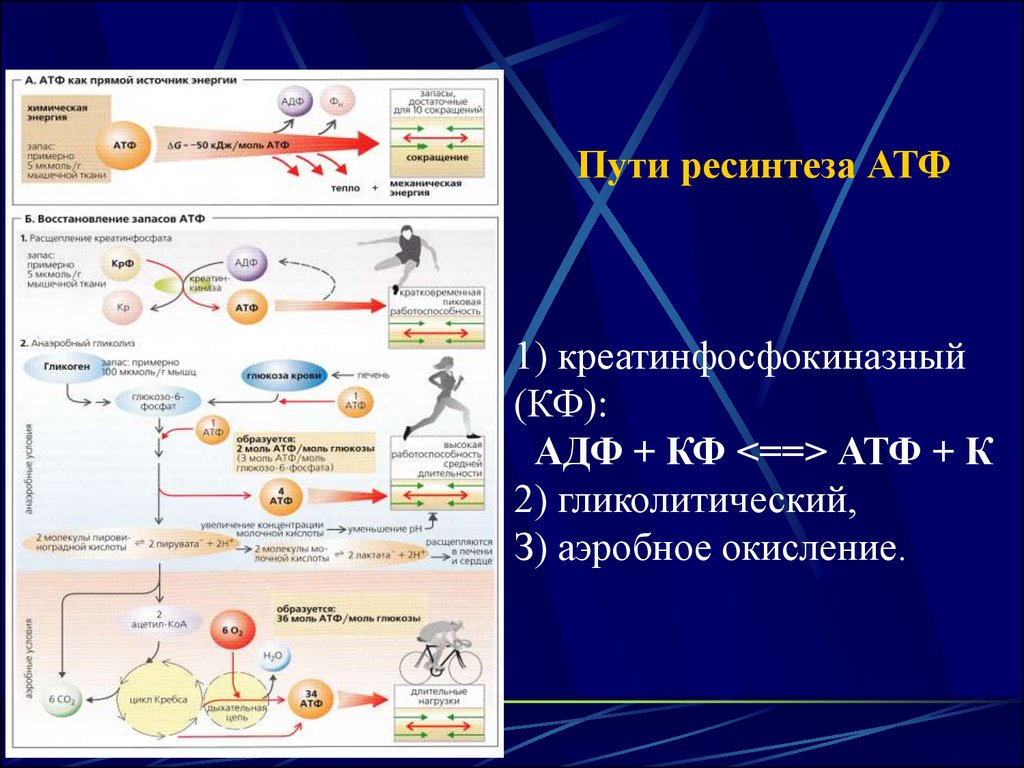
Пути ресинтеза АТФ1) креатинфосфокиназный
(КФ):
АДФ + КФ <==> АТФ + К
2) гликолитический,
З) аэробное окисление.
28. Максимальная мощность различных путей ресинтеза АТФ
а) фосфагенный (КФ) - 3,6 мольАТФ/мин,
б) гликолитический - 1,2 моль АТФ/мин,
в) окислительный - при окислении
глюкозы - 0,8 моль/мин, жиров - 0,4
моль/мин.
29. Двигательная единица - единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна, составляют одну ДЕ
1 - тело мотонейрона;2 - ядро;
3 - дендриты;
4 - аксон;
5 - миелиновая
оболочка аксона;
6 - концевые веточки
аксона;
7 - нервно-мышечные
синапсы.
30. Быстрые и медленные ДЕ
БыстрыеБольшой мотонейрон
(высокая скорость
проведения ПД).
Много АТФ.
Много КФ.
Активный гликолиз.
Медленные
Малый мотонейрон.
Меньше АТФ и КФ.
Менее активный
гликолиз.
Много митохондрий
(активное окисление).
Сильные, но быстро
устающие.
Способны выполнять
длительную работу.